CHAPTER I: WHAT PLANTS ARE
WHAT we commonly call plants, such as corn or buttercups or an oak tree, are so familiar that a definition of what plants are may seem needless. It would be quite so if these generally recognized examples comprised all the plant kingdom. Actually what are ordinarily thought of as plants make up only a fraction of the great plant world. The fact that our familiar roadside and garden plants produce blossoms followed by a fruit and seed, such as peas and beans and all the ordinary flora of any region, separates them at once from that other large group of plants that do not. Common examples of the latter class are the green scum on the ponds, moss, seaweed, the rust on wheat, yeast, disease-causing bacteria, the smallest of all known plants, and many others. Most of these organisms are so small that they can be distinguished only under the higher powers of the microscope. Some of them in their habits and growth are like the minute animals described in the volume of this series devoted to that subject. In fact there are organisms about which scientists are still in doubt as to their animal or vegetable character.
One or two characteristics common to most plants, however, separate them from animals and these are their method of getting food and their practically immovable mode of life. Animals, however simple, do eat and digest their food, plants take various mineral substances from the earth or air or water in the form of inorganic matter such as oxygen, carbon, nitrogen, and all the food materials found in the soil, and transform them, with the aid of sunshine, into the structure that characterizes each particular form. Plants, then, may be defined as any living organism that, with minor exceptions, has the power to assimilate inorganic substances and transform them into organic matter. Nothing else in all the realm of nature has this power. It is a possession beyond all price, this ability to take from the soil and air and water simple chemical substances and under the magic of sunlight transform them into the wonderful plant life of the world. But this faculty has its handicaps, for it is usually, though not always, associated with inability to move from place to place, which, in some measure, even the simplest animals can accomplish.
It will be readily understood that this definition of plants includes many other things that are commonly attributed to the plant kingdom. For our purpose the discussion of these relatively unfamiliar forms of plant life will be left until later. A rough and ready distinction between those plants that everyone recognizes as such and those others like yeast and microbes, which are plants to most of us only by virtue of the definition given above, is that the first group produce flowers and seeds and the second do not. It should not be forgotten, and it will be shown later, that this is not a true distinction, but for practical purposes of dividing the plant kingdom it suffices.
1. Flowering Plants
The fact of outstanding importance to everyone who really looks at most plants is that part of them are above ground and part below. This simple observation carries with it the recognition of a fundamental difference of plant structure, namely roots and stem. Most plants bear obvious leaves, and at some time in their life flowers, inevitably followed by fruits and seeds. The ideally perfect plant would consist, then, of root, stem, leaves, flowers, fruit, and seed. These are subject to many changes of form, sometimes they are put to strange uses, and occasionally one or two may be lacking, as the stem is apparently from many violets, and leaves from some cactus and from the Tjemoro tree of Malaya. In fact, so varied are the different forms of these common attributes of most plants, so important are these differences of structure that no right understanding of plant life can be had without examining each in some detail.
THE ROOT
The obvious purpose of the roots of plants is to serve as an anchorage or holdfast. Their other and equally important function is to secure food for the plant, a process that will be described in the part devoted to Plant Behavior. Certain plants bear no roots and attach themselves to the roots of other plants in which case they literally steal their food, as does the mistletoe and some others.
Roots are of various kinds, depending upon the soil in which they grow and upon the kind of plant to which they are attached. In the case of annuals, which live only one year, as does the purslane, and biennials, which live only two years, as does the fringed gentian, the roots are mostly fibrous (Figure 1) and apt to be only slightly under the surface. In perennials, which live many years, such as the dandelion, the root is deeper and forms what is known as a taproot (Figure 2). In shrubs and trees they are harder, woodier, and often penetrate to great depths.
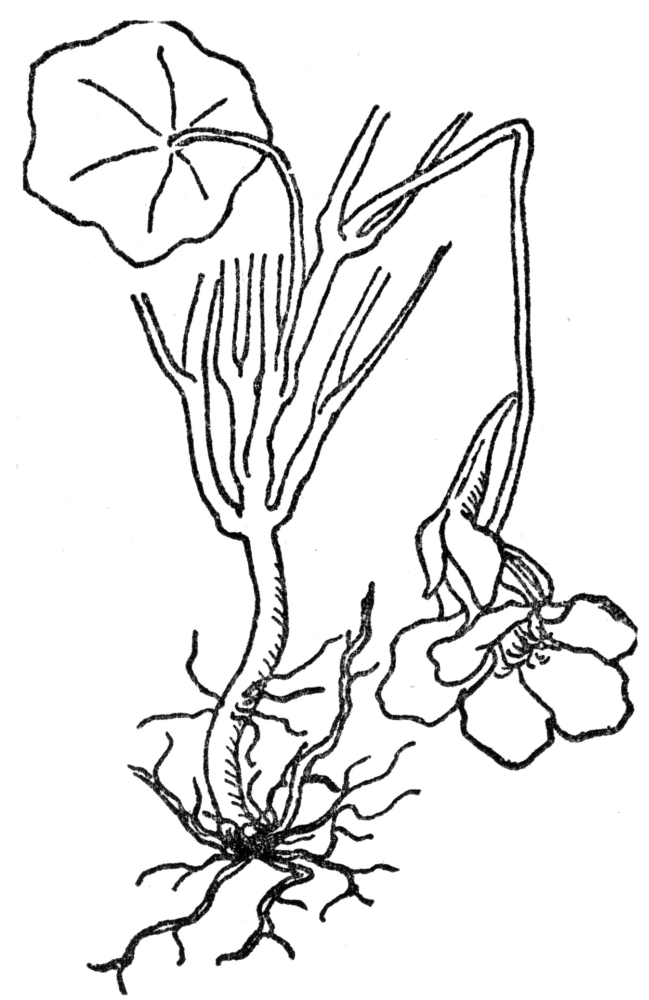 FIG. 1.—FIBROUS ROOTS As illustrated by the common garden nasturtium.
FIG. 1.—FIBROUS ROOTS As illustrated by the common garden nasturtium.
If we examine the roots of a tree, we find a large part of them are woody, often as thick as the smaller branches, and it is only toward their extremities that they branch out into the multitude of rootlets
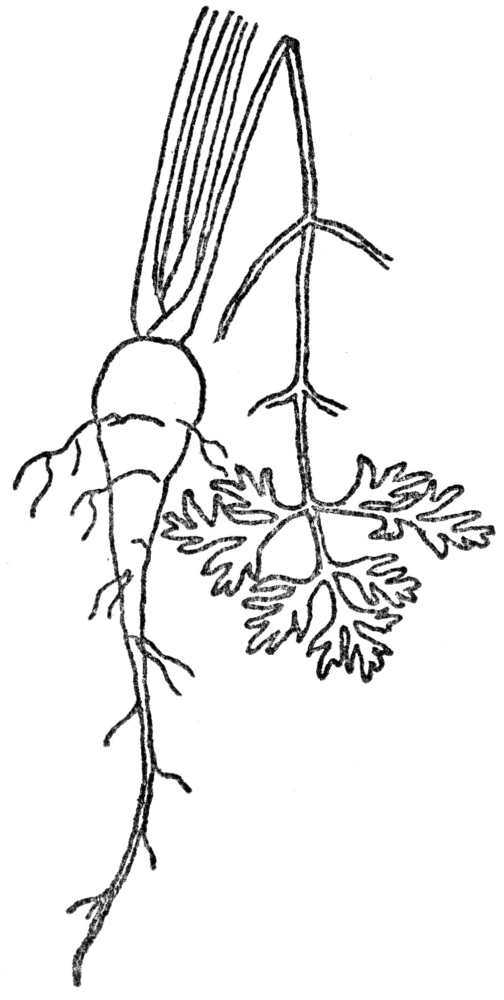 FIG. 2.—TAP ROOT OF CARROT A store of food for the second year.
FIG. 2.—TAP ROOT OF CARROT A store of food for the second year.
that creep through the earth seeking food for the tree. Some, as in the spruce or hemlock, do not go very deep but spread great distances through the soil in search of food, others, like the hickory, go nearly straight down. The interesting feature of these tree roots is that the part nearest the trunk is all but dead, and acts mostly as an anchor, while the fibrous rootlets or even finer subdivisions known as root hairs at the extremities are the food gatherers. At the very end of all rootlets and of roots is a rootcap (Figure 3), harder than the rest of the threadlike rootlet. This rootcap is, if not quite dead, at least useless as a food gatherer, but serves as a tiny pioneer wedge which forces its way among stones or other obstructions, so allowing the living root hairs just behind to gather the food to which it leads the way. In certain of the rocky islands of the Bahamas wild fig trees may be seen, growing on the bare rocks, their roots sprawling in every direction in search of a crevice through which the rootcap can force its way. Such roots may extend thirty or forty feet from the trunk of the tree over the bare rock in search of a favorable crack where they plunge to the cool depths and secure the food and water necessary for life.
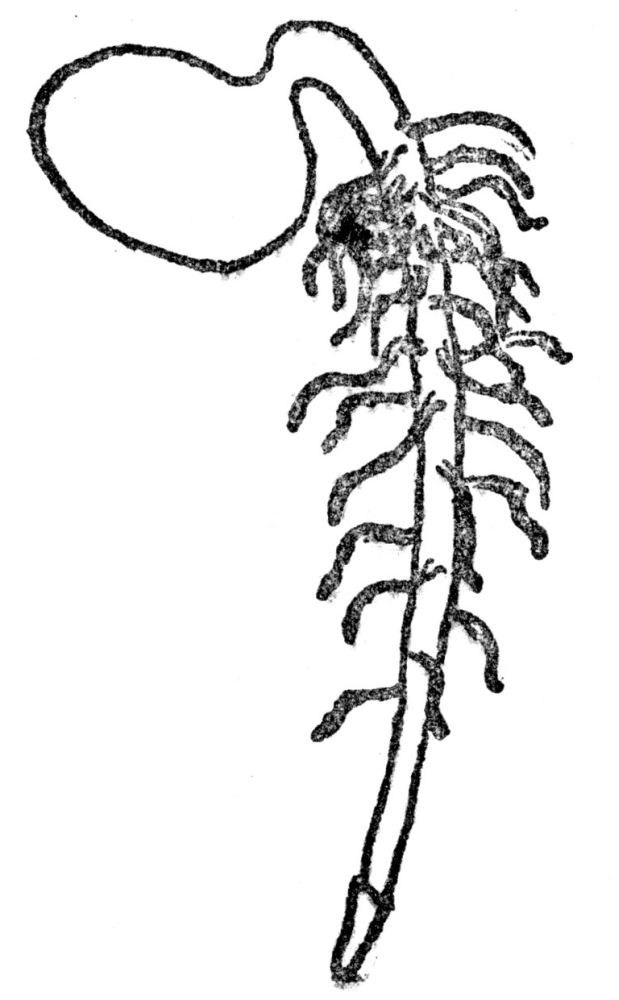 FIG. 3.—TAP ROOT OF SEEDLING The figure shows the root cap at the extremity.
FIG. 3.—TAP ROOT OF SEEDLING The figure shows the root cap at the extremity.
Roots are not always of this common type. Sometimes, particularly in certain biennials, they are swelled to form great thickened portions, often weighing many pounds. The sweet potato is a familiar example, and a related plant, one of our morning-glories, has an enormous perennial root, known to weigh as much as fifty pounds. This swelling of the roots of plants is a quite common characteristic of certain kinds and has great commercial significance. Carrots, turnips, rutabagas, beets, and parsnips are familiar examples. The purpose of such roots is to store food for the plant, and this thrifty habit of some roots has been turned by the gardeners to our advantage.
It is a common sight to see parts of a sidewalk heaved up by a tree root and their force in this respect is tremendous. One of our common ferns has been known to raise a weight of over 500 pounds, and even to break through a concrete walk. Such is the force exerted by the roots of plants that we can truly think of roots as pushing through the earth almost regardless of obstacles, binding the soil together and not only serving the needs of the plants, but actually holding the soil on steep slopes. Where fire or ruthless lumbering has stopped this natural process the washing away of the soil and exposure of the bare rocks leaves desolation behind it.
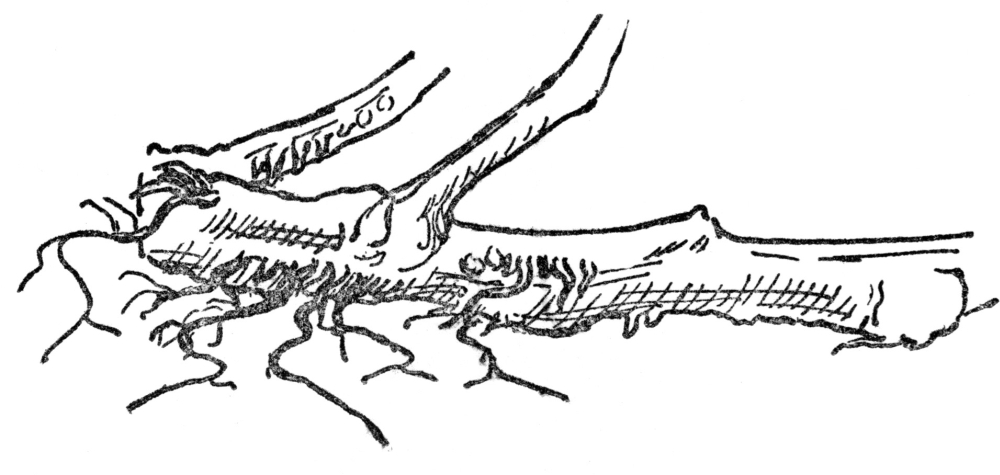 FIG. 4.—ADVENTITIOUS ROOTS OF THE TOMATO Produced from joints or injured places.
FIG. 4.—ADVENTITIOUS ROOTS OF THE TOMATO Produced from joints or injured places.
While most roots live under the surface many grow in the air, and a few grow from stems that are injured. The tomato vine often produces roots at the joints or where it has been injured. Such roots, known as adventitious roots (Figures 4 and 5), are fairly common in many plants, the common garden practice of making cuttings, which take root under favorable conditions, being based upon this fact.
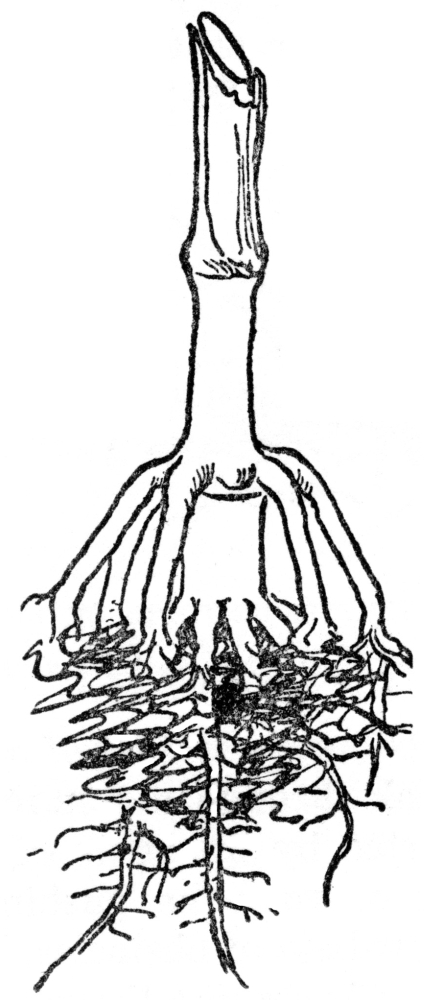 FIG. 5.—PROP ROOTS OF THE INDIAN CORN Usually produced from the first joint above ground and serving as additional anchor and food gatherers.
FIG. 5.—PROP ROOTS OF THE INDIAN CORN Usually produced from the first joint above ground and serving as additional anchor and food gatherers.
But some plants produce roots in the air, as in poison ivy and the trumpet creeper, without injury or the gardener’s skill, and are known as aërial roots. They are some of the most peculiar and fantastic of nature’s devices for allowing plants to grow in apparently unfavorable places. In many orchids, some relatives of the pineapple, and a few other air-inhabiting plants, the roots live wholly in the air, the plants being fastened to a tree or even to a telegraph wire. Such plants live on the air and water vapor, and are mostly inhabitants of moist tropical regions.
Quite the most extraordinary of aërial roots are those produced in certain kinds of fig trees. Starting perhaps a hundred feet in the air and no thicker than a lead pencil they appear first as slender vine-like streamers blown hither and yon by the wind. Eventually they reach the ground and penetrate it, grow often to a great size and even form trunklike connections with the tree top. The banyan tree of India is the best known case of this peculiar habit. One which started from a single trunk, subsequently sending out great numbers of aërial roots, has now spread to such a size that it is over 2,000 feet in circumference, has 3,000 trunks, and once sheltered 7,000 soldiers.
A variation of this habit is the case of a giant fig tree of the West Indies in which a bird may deposit the seed of another tree. The seedling soon develops, sending out long, at first threadlike, aërial roots which are wrapped around the tree trunk. As the roots increase in size and further encircle the trunk they ultimately reach the ground, where they are frequently a foot in diameter. Then the true nature of the process becomes evident. For these apparently innocent aërial roots, as they reach the ground, have so completely inclosed the old trunk and their pressure is so great that they literally strangle the tree from which they started. It is slower but just as deadly as the strangulation of an animal by a boa constrictor, for these encircling roots cut off by strangulation the ascent of the sap, thus killing the tree. Fate sometimes overtakes them, however, as it is a common sight to see the strangler meet the same end. Some of nature’s most ruthless battles are fought out in this way, very silently, but very effectively.
THE STEM
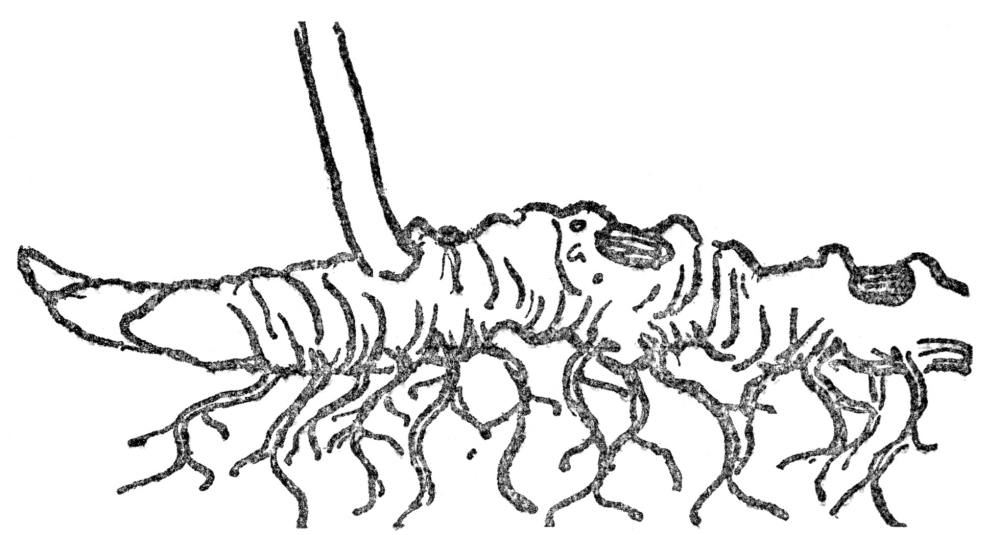 FIG. 6.—ROOTSTOCK OF SOLOMON’S SEAL The figure shows roots and bud scars together with the ascending stem of the year’s growth.
FIG. 6.—ROOTSTOCK OF SOLOMON’S SEAL The figure shows roots and bud scars together with the ascending stem of the year’s growth.
Nothing dies harder than generally accepted delusions, particularly those regarding plant lore, and of all such incorrect impressions the one that a potato is a root, is the hardiest and most difficult to kill. Yet, the “eyes” of a potato give it away if one stops for a moment to reflect that the eyes are only buds and buds grow only on stems. That is one of the chief uses of stems—to support in the air the leaves and flowers that come from its buds, and no matter if the stem, as in the potato and many other plants, be ever so deeply buried their true stem nature cannot be mistaken. Sometimes these underground stems are not thickened but lengthened out, in which case, notably in common garden iris, they are called rootstocks. Again, these buried stems may be swollen, as in the potato, when they are known as tubers. Onions and the jack-in-the-pulpit bear still other kinds of underground stems, and there are many more, but they cannot be mistaken for roots, for it will be seen from Figures 6-9 that on their under sides they bear roots themselves. Besides this they bear buds or shoots, which no true root ever does.
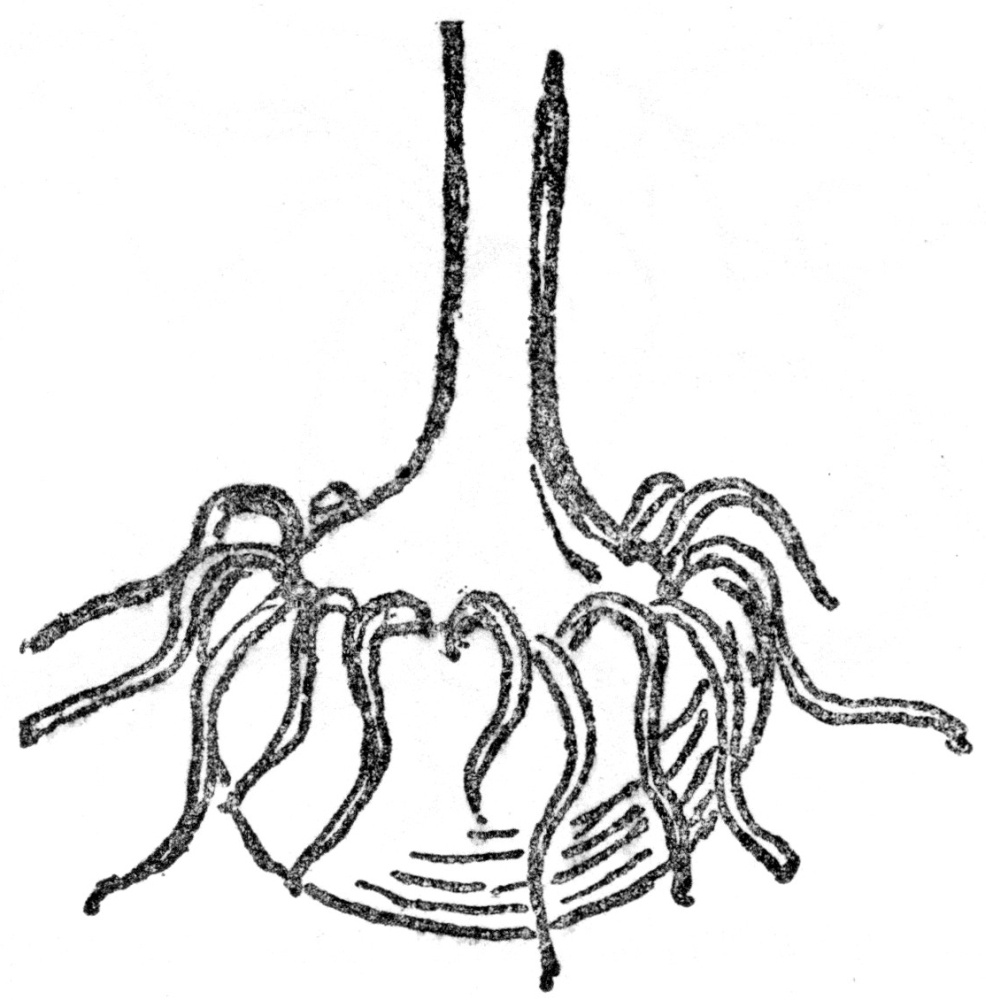 Fig. 7.—CORM OF JACK-IN-THE PULPIT (After Gray) Really an underground stem.
Fig. 7.—CORM OF JACK-IN-THE PULPIT (After Gray) Really an underground stem.
Stems above ground, which is the most usual form for them, are of many kinds, all serving the purpose of support to the leaves and flowers, and as a means of carrying sap from the roots or underground stems to the upper part of the plant, and also to carry certain foods to the roots from the leaves, of which more anon. In the case of herbs, like goldenrod or daisy, the stem may be apparently all pith on the inside, with only a thin outer coating of harder substance, not unlike bark, but usually green. If we examine the cut-off trunk of a tree, a quite
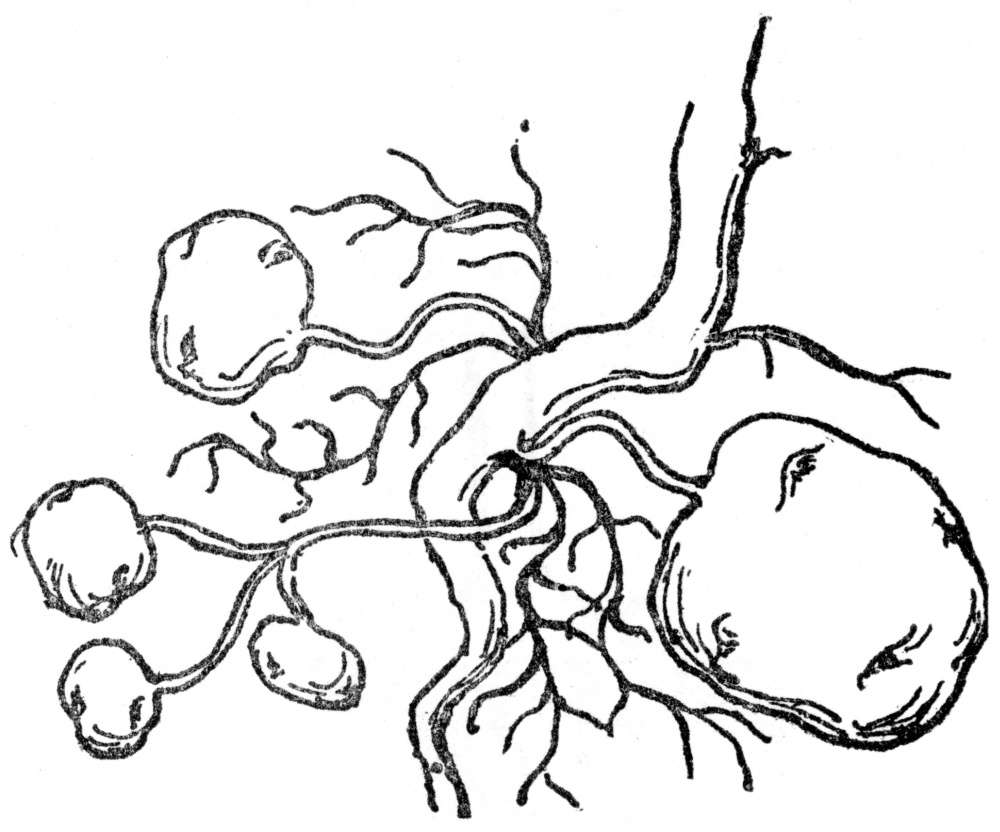 FIG. 8.—TUBERS OF THE COMMON POTATO Potatoes are swollen portions of rootstock.
FIG. 8.—TUBERS OF THE COMMON POTATO Potatoes are swollen portions of rootstock.
different structure is apparent. Any lumberman can point out at once “heartwood” and “sapwood” (Figure 78), and his distinctions are just as good as those of the scientist, for he says in these two words as plainly as can be said that heartwood is the oldest and sapwood the youngest. The sapwood is nearer the bark and is honeycombed with passages which serve to carry the sap from the roots to the tree top, while just under the bark is the bright, green, living layer, known as cambium, which is renewed each year. The phloem is the carrier for the food made in the leaves to the roots. It is the successive layers of cambium, year after year, that gives to tree trunks their annual rings. The age of almost all trees can be reckoned exactly by counting these, one representing a year’s growth, and the tree’s rate of growth estimated from the closeness of the rings. Fires or droughts, perhaps long forgotten, here find a lasting record in rings so close together as to be all but invisible. The part nearest the center of the trunk is the heartwood, usually quite lifeless, yet in its maturity furnishing us with lumber. It may be and often is completely decayed, without injuring the flow of sap or the life of the tree for many years.
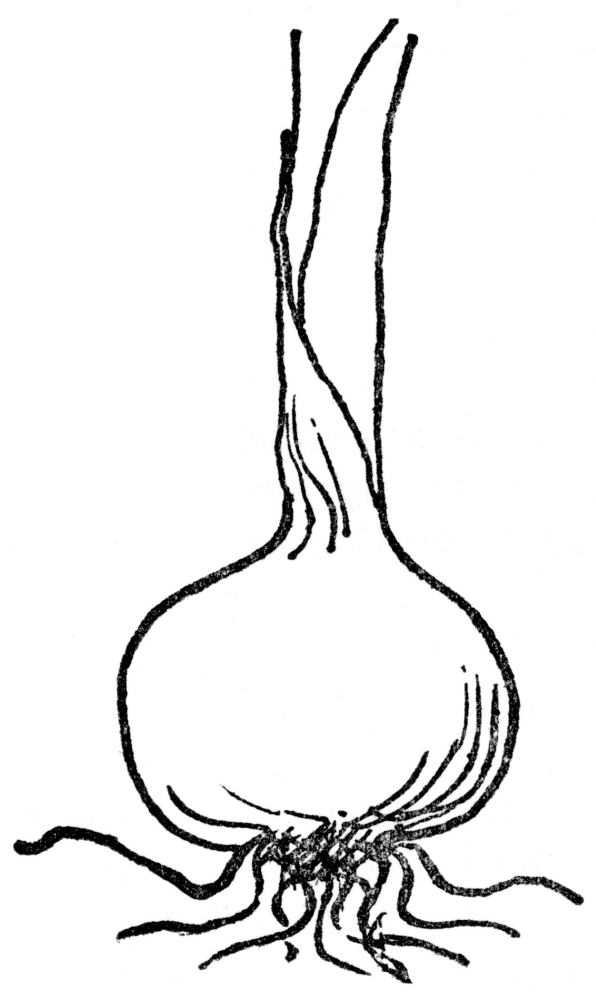 FIG. 9.—BULB OF THE ONION Showing root and leaf growth.
FIG. 9.—BULB OF THE ONION Showing root and leaf growth.
These two streams of sap, one going up and the other returning to the roots, each in its proper channel, are interspersed with air chambers that extend from the center of the tree out toward the bark, where they end in inconspicuous dots called lenticels. It is as though nature had provided an air-cooling device for the constant activity of these diverse currents. These lenticels are prominent on the bark of cherry, but whether obvious or not they are found in nearly all woody stems and insure a constant supply of fresh air to the busy interior.
In palms, sugar cane, corn, bamboo, and many other plants there is not any distinction between heartwood and sapwood (Figure 82), and in place of bark there is nothing but an outer rind, harder than the interior tissue. Such stems do not usually rot first at the center, have no cambium, and have no annual rings. This method of growth and structure is associated nearly always with definite leaf and flower forms peculiar to it and differing from most other plants. So fundamental are these characteristics, so uniform their occurrence and so clear are the distinctions between them and other plants that botanists have divided all flowering plants into those belonging to this group or to some others. More will be said of this in the chapter on the Families of Plants and Their Relationship.
The stems of some plants, such as the Big Trees of California, for instance, are among the oldest and most permanent of living things. “General Sherman,” one of the biggest in that most famous grove, was nearly three thousand five hundred years old when Columbus discovered America; it has lived through all the great periods of modern history, and to-day it is over 270 feet high and 35 feet in diameter. No living thing is so large or has lived so long. In Australia are great forests of blue gum trees even taller than our Californian Big Trees, but not so old nor so thick.
In the Pacific, off the coast of Oregon and British Columbia, a seaweed is commonly found with stalks over 500 feet long, and in India the rattan palm climbs over the tree tops for great distances, a single stem not much thicker than a broomstick measuring over 700 feet long. The search by leaves for light and air results in the stems of some plants performing almost incredible feats. Whether it is one of the Big Trees with a great massive trunk, or the rattan palm with its sinuous winding through the topmost heights of the tropical forests of India, the result is always upward to a “place in the sun.” This struggle for sunlight has taken many forms in different plants, the ordinary vines like morning-glory or grapevine, for instance, where the climbing stem is of great advantage. Some vines always twine to the left, as the hopvine, others to the right, as in the morning-glory, all seeking support from something else, each adopting its own most useful way of getting its leaves in the most advantageous position to catch the life-giving sunshine. If we could look down on any forest from an aeroplane, the striking efforts of nearly all plants, whether herbs, shrubs, vines, or trees, to get the utmost sunshine for their leaves would be evident at once. No apparently impossible twisting or bending of tree trunks or reaching out of stems of vines but is to be found in the inexorable struggle of stems to fulfill their task of giving the plant its chance to reach “a place in the sun.” Sometimes mere climbing or twining does not seem sure enough—it seems as though winds or the elements might break loose the vine from its support and thereby kill its chances. In certain vines this contingency appears to have been foreseen, and as if to clinch their opportunity of growing onward they are provided with special helps. Slender green tendrils, delicate prolongations of the stem, begin, almost insidiously, to catch hold of the nearest support and by a couple of turns about it and subsequent strengthening of their tissues make a permanent holdfast. The grapevine is a case in point. And as if this were not enough, certain other plants, such as the Boston ivy, have small disks which attach themselves to bare walls or tree trunks. This is to make assurance doubly sure, and it is this that makes the Boston ivy so useful to the gardeners for covering walls.
Some stems accomplish their purpose not by holding fast to a support in the air, but by creeping along the ground, as in the running blackberry, and often in the Virginia creeper. The purpose is the same, and, as if to confirm it, a few otherwise quite prostrate vines have their tips turned upward to the light, notably in the case of the creeping speedwell.
In certain plants the stem may assume curious forms due to special conditions under which they live and to which adjustment is necessary for the plant’s existence. In deserts, for instance, the cactus produces practically no leaves (Figure 10) and the green stem performs not only the function of leaves but acts as a storage for water. Where water is scarce this is of tremendous advantage, a single cactus having been known to store up 125 gallons. A similar habit of the cactuslike spurges in South Africa gives as weird an atmosphere to parts of their landscapes as we find in Arizona. It is as if the stem of such plants, being unable to push its leaves (it has none) up into the light, takes over some of the functions of leaves and makes up the deficiency by adopting other methods
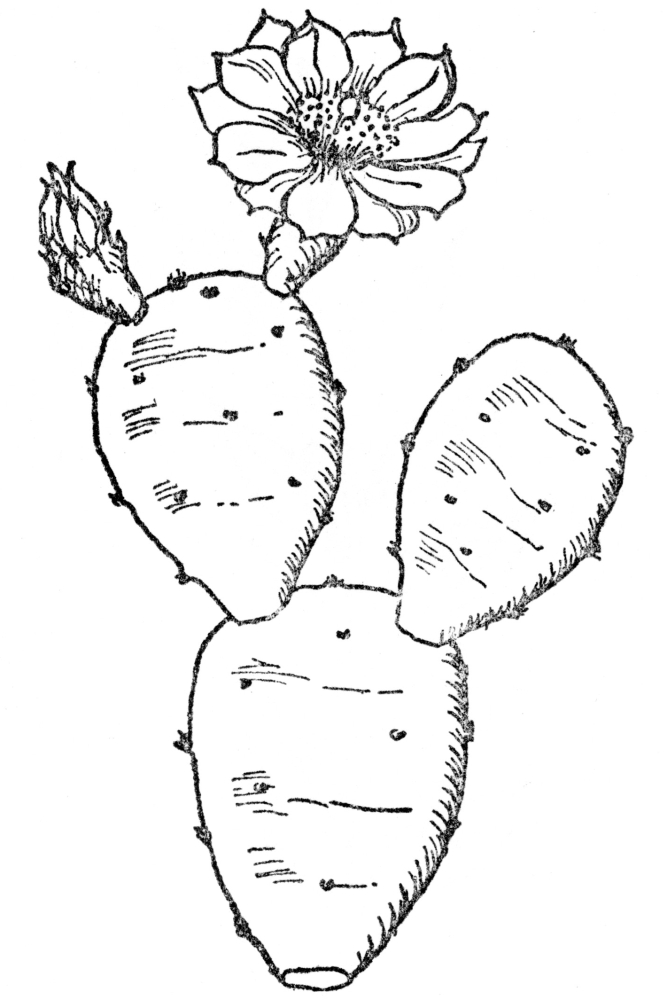 FIG. 10.—COMMON PRICKLY PEAR CACTUS (Opuntia Opuntia) Native along the Atlantic Coast. The green joints of the stem function as leaves and store water.
FIG. 10.—COMMON PRICKLY PEAR CACTUS (Opuntia Opuntia) Native along the Atlantic Coast. The green joints of the stem function as leaves and store water.
to secure the plant’s survival. Other stems, looking and acting like leaves, reveal their true nature by producing buds, and the curious feature of the common butcher’s-broom (Figure 11), often colored scarlet for Christmas decorations, bearing flowers from the middle of what is apparently a leaf, but is actually a modified stem, is explained by this ability of stems to modify their habits to suit conditions. The butcher’s-broom is an inhabitant of dry regions along the Mediterranean, where a reduction or
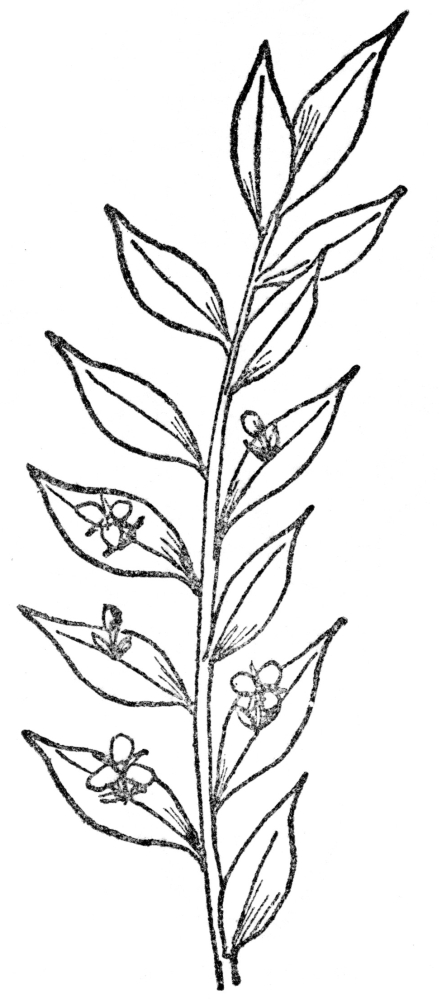 FIG. 11.—BUTCHER’S-BROOM (Ruscus aculeatus) Note leaflike stems with flowers arising from the center.
FIG. 11.—BUTCHER’S-BROOM (Ruscus aculeatus) Note leaflike stems with flowers arising from the center.
absence of leaf surface is a decided advantage. In many partly desert or dry regions this production of leaflike stems or branches is common, an excellent garden example being asparagus, which came originally from Europe and the feathery growth of which is all stem. In Tasmania a kind of yew tree produces no leaves, all the foliage being modified stem, which is true of many kinds of spurge in the West Indies, where an almost impenetrable scrub is largely made up of a shrub which is apparently covered with leaves, all actually part of the branches and stems.
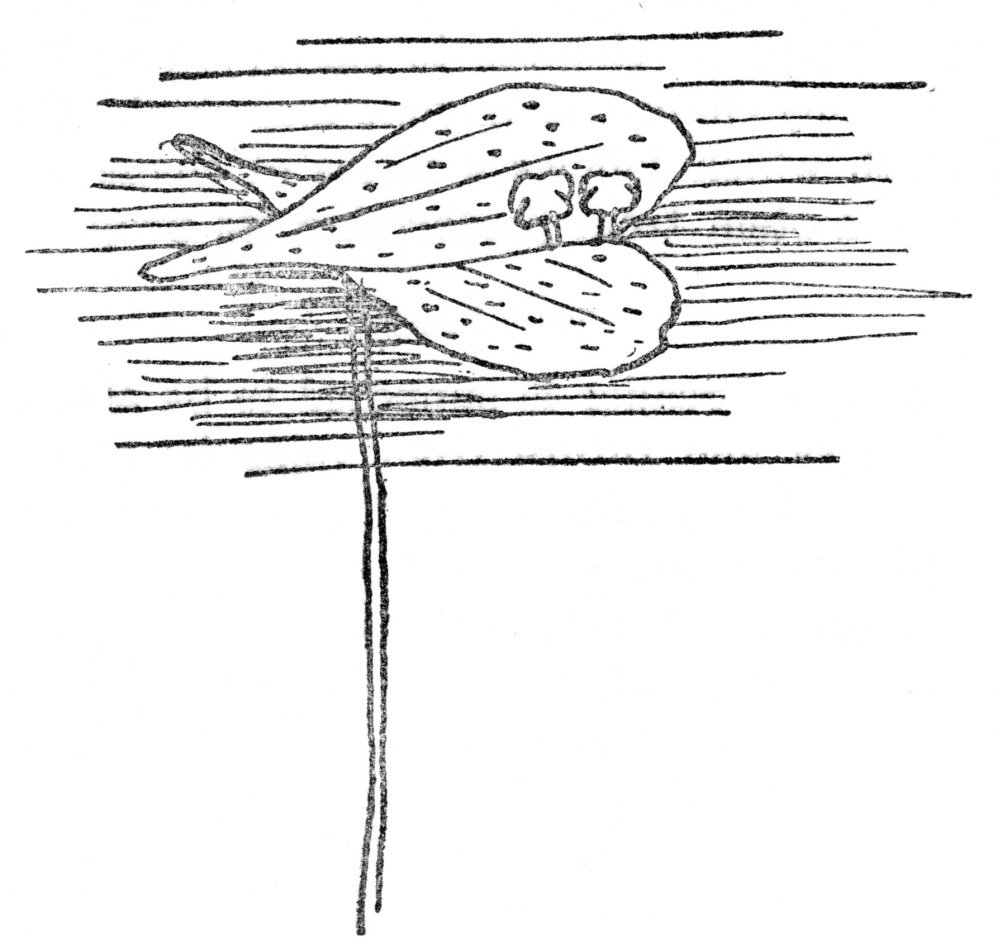 FIG. 12.—DUCKWEED The smallest known flowering plant, with no leaves and tiny leaflike stems floating on the surface. Flowers are borne from the margin of the stem. (Eight times natural size).
FIG. 12.—DUCKWEED The smallest known flowering plant, with no leaves and tiny leaflike stems floating on the surface. Flowers are borne from the margin of the stem. (Eight times natural size).
While stems, such as the Big Trees or the giant cactus, may be among the largest of nature’s creations they may be also the smallest, as the duckweed that floats on ponds is the tiniest of all flowering plants and its flat expanded surface is wholly stem. Figure 12 on this page better illustrates this strange modification of a stem than words could do.
From what has been read it will be seen that stems are not “just stems”—they are among nature’s most ingenious devices to secure the survival of the plant. Whether buried in the ground, and producing, almost by stealth, buds that develop into mature plants, or thrusting leaves to the utmost limits of their reach, or climbing by an intricately varied mechanism, or changing their character to suit desert conditions, or floating on the water—it matters not. Each modification of form or use secures to the individual plant its chances to survive; and in most cases its only chance, as anyone may see by the sudden death which follows a series of changes which prevents a stem from performing its proper tasks.
THE LEAF
As the palm reader is supposed to be able to tell your history and future from veins in your hand, and as the veins in the wing of a butterfly tell their story to an entomologist, so the veins of a leaf are more significant than almost any other characteristic of a plant. Most leaves have their veins, or skeleton, with a single midrib and many branches off it on each side, which themselves break up into a fine network of veins. Such leaves are netveined (Figures 13-24). Others, such as corn and grass, have the veins running side by side from one end of the leaf to the other, sometimes with small branches off them, but instead of the veins forming a network they are parallel, and such are called parallel-veined leaves (Figure 38). In the chapter on Plant Families and Their Relationship more will be said as to the amazing regularity with which netveined leaves are associated with certain kinds of flowers and parallel-veined with other kinds, how these distinctions have been recognized since hundreds of years before Christ, long before their true import was understood. There are variations from both these

Banyan Tree (Ficus bengalensis). A fig tree of India, whose adventitious roots make frequent connection between the tree top and the ground. Starting as thin, whiplike streamers these roots ultimately form new trunks. (Courtesy Brooklyn Botanic Garden.)

Laceleaf (Aponogeton fenestralis). A submerged aquatic plant, with permanently skeletonized leaves, and an inhabitant of forest pools in Madagascar. (After Engler & Prantl. Courtesy of Brooklyn Botanic Garden.)
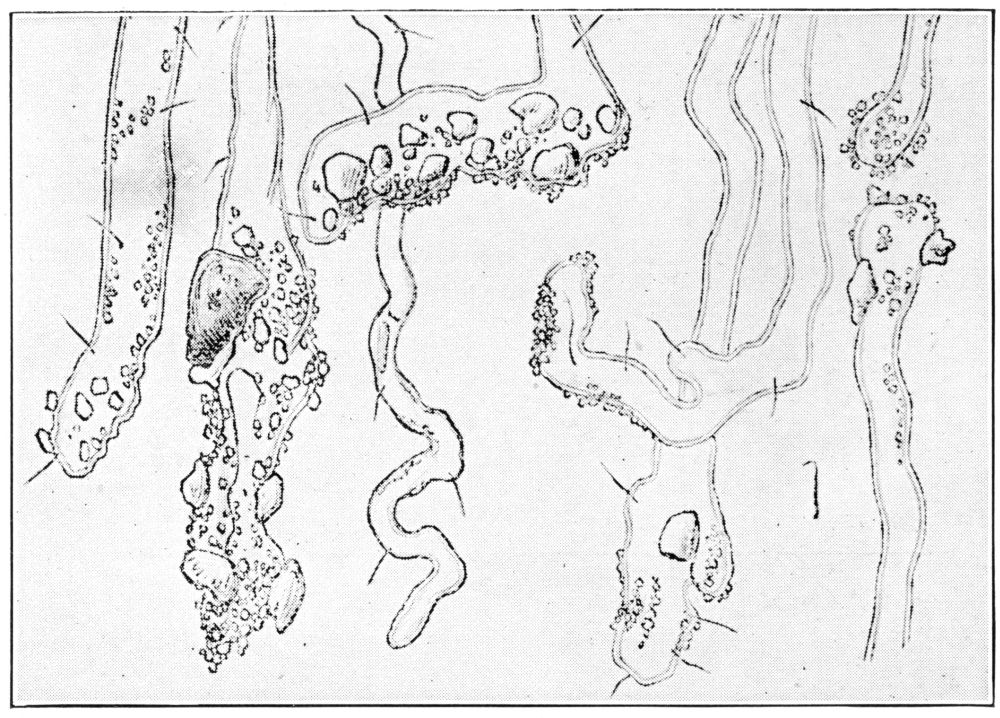
Root Hairs with Fine Soil Particles Attached. Note soil, water, and air spaces. Much magnified. (After U. S. Department of Agriculture. Courtesy of Brooklyn Botanic Garden.)
 FIG. 13-24.—FORMS AND TIPS OF LEAVES
FIG. 13-24.—FORMS AND TIPS OF LEAVES
Fig. 13. A linear leaf with an acute tip. Fig. 14. Lanceolate leaf with an acuminate tip. Fig. 15. Oblanceolate leaf broadest above the middle. Fig. 16. Ovate, broadest below the middle. Fig. 17. Spatulate, broadest above the middle and with an elongated base. Fig. 18. Elliptical. Fig. 19. Obovate in which the general shape is ovate, but broadest toward the tip. Fig. 20. Oblong. Fig. 21. Orbicular or nearly round. Fig. 22. Deltoid or somewhat triangular, an ovate leaf with a broad base. Fig. 23. Kidney-shaped or reniform with heart-shaped base. Fig. 24. Peltate leaf of common garden nasturtium; note circular blade with leafstalk attached to the center.
types, but in nearly every case, once the difference is noted—and scarcely any other character of a plant is so much worth notice—they cannot be mistaken.
During the winter nearly all leaves are folded in various ways in a bud for protection from the elements. Nature shows herself in some of her wisest moods in the selection of methods to accomplish this. In some buds, notably those of the horse-chestnut, the bud is coated with a sticky substance to protect the tender young leaves inside. In others there is a hard outer coat, as in the hickory, impregnable to the most driving sleet, others again have the leaf rolled so tightly and pointed so sharply at the end, as in the beech, that water cannot cling to the bud nor soak in, until the warmth of spring gives the signal for the annual miracle of the bursting out of foliage. Leaf buds are sometimes hard to find on certain plants, as they are formed at the base of a leafstalk and covered by it during the growing season. It is only as the leaf falls in the autumn that the hollow base of its stalk is seen to have hidden during the summer the young bud for the following season. The plane tree or sycamore is a good example of a plant where no leaf buds can be found until the falling of the leaves in autumn.
The forms of leaves are infinite in their variety, and the reasons for some of their peculiarities in this respect are not yet understood. The average netveined leaf is obviously composed of a blade (Figure 25), and at the base a stalk known as a petiole. Sometimes at the base of the petiole—which is lacking in many leaves—there are two tiny leaflike appendages, called stipules, which are of no apparent use to the plant, and, as if in recognition of this fact, they often fall off long before autumn. In some plants, however, stipules are permanent, while in certain others they are never found, as, for instance, in the horse-chestnut tree.
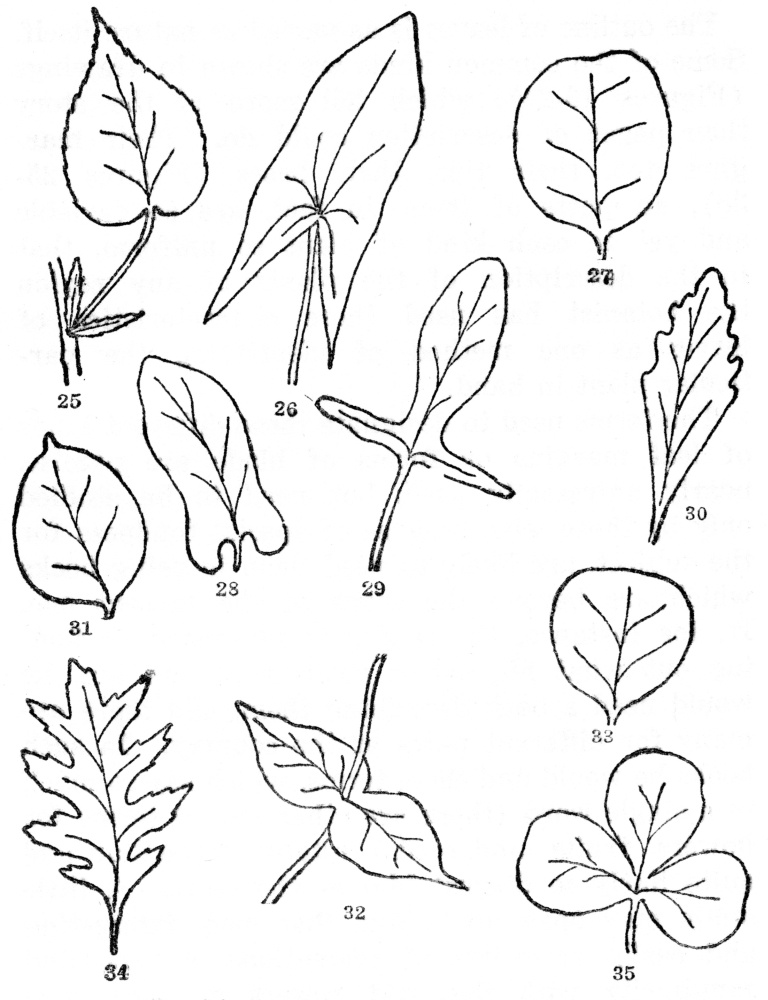 FIG. 25-35.—FORMS AND BASES OF LEAVES
FIG. 25-35.—FORMS AND BASES OF LEAVES
Fig. 25. Simple leaf with blade, leafstalk (petiole), and two stipules at the base. Margins of the leafblade serrate or saw-toothed. Fig. 26. Leaf with a sagittate base, or shaped like an arrowhead, the lobes pointing downward, and with entire margins. Fig. 27. Retuse or emarginate tip, somewhat indented. Fig. 28. With the base auriculate or with rounded basal lobes. Fig. 29. Hastate, like an arrowhead but the lobes pointing outward. Fig. 30. With cuneate base (wedge-shaped). Fig. 31. Cuspidate tip with a usually hard and stiff point. Fig. 32. Perfoliate, the leaf bases joined and the stem passing through them. Fig. 33. Truncate, the top flattened. Fig. 34. Pinnately lobed, with deep indentations cut toward the midrib. Fig. 35. Palmately lobed, out toward the top of the leafstalk.
The outline of leaves is as varied as nature itself. Some of the common kinds are shown in drawings (Figures 13-24), which tell more of the story than pages of description could do. Their margins, too, their tips, their bases (Figures 25-35), all parts of them, in fact, are so variable and yet in each kind of plant so uniform, that in the description of the plants of any region the botanist has used these characteristics of leaves as one method of identifying the particular plant in hand.
The terms used to designate these different kinds of leaf margins or forms of blade are precise, nearly universally used, but need to be studied only by those who, because of special fondness for the subject, are likely to need them in using books which are beyond the scope of the present one. If, for instance, the reader is interested in finding out what his native roadside plants are, he would need a book describing them, and there are many for different parts of the country. In such books he would find these terms, which say so much in a single word (there are other sets of terms for flowers, fruits, and seeds) totally unfamiliar and quite likely to disgust him at the start. A little study may open up to him that most interesting and easily accessible of recreations, a first-hand familiarity with the wild flowers of one’s own neighborhood.
All leaves are not as simple as the figures show them to be. In many the midrib or principal vein is much elongated and there are small leaflets, sometimes even scores of them, all fastened to a common stalk. Such are called compound leaves (Figures 36-37), which may be found in ash, hickory, rosebushes, blackberries, peas, beans, and thousands of other plants.
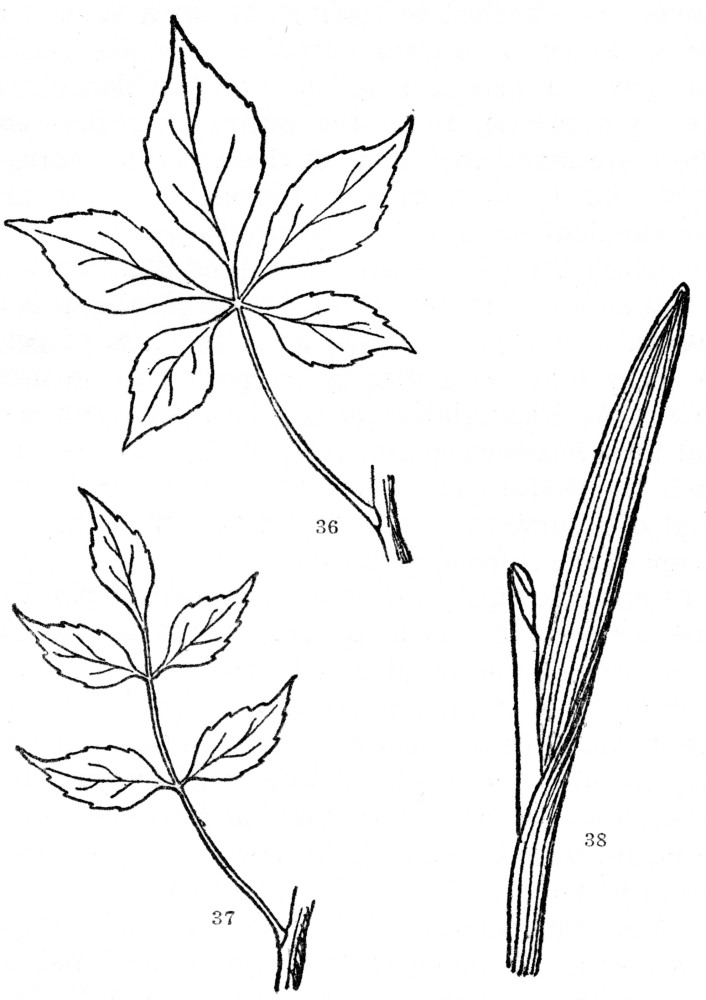 FIG. 36-38.—COMPOUND AND PARALLEL-VEINED LEAVES
FIG. 36-38.—COMPOUND AND PARALLEL-VEINED LEAVES
Fig. 36. Palmately compound leaf, the five leaflets all arising from the tip of the common leafstalk. Fig. 37. Pinnately compound leaf, the leaflets arising from the sides of the common leafstalk. Fig 38. A parallel-veined leaf. All the other leaves figured are netted-veined.
While leaves are literally factories in which one of the most wonderful things in the world is produced, it is so much a part of what plants do or their behavior that the story of it will be given in the chapter on Plant Behavior. Sunlight is absolutely necessary for the process, and to reach this sunlight leaves are attached to their stems in a variety of ways. Some are always opposite each other, as in the common privet, lilac, or honeysuckle; others always alternate, as in the mustard or the rose. There are many variations of these simple arrangements, but in every case the process results in giving each leaf the utmost exposure to the light without which the plant must wither and die. So vital is this exposure to light that in some plants parts of the leaves produce tendrils, as in the case of peas, in order that some near-by support may be used. In one African relative of our lily, this change of leaf form has been so great that its long slender leaf tip is wonderfully adapted to reaching up and catching by its curved tip some support to lift it from the gloom of the tropical forest floor.
Looking down from above on any small plant or bush, or from the sky on a forest, about all that can be seen are the thousands of leaves, all so arranged that it is as though some celestial photographer asked every one of them to so place themselves that they would all be “in the picture.” The competition between leaves on the same plant and between leaves on rival plants is infinitely keener than the friendly pushing of a crowd to get in a picture, and it lasts forever. Furthermore, failure to get in means certain death. So intricate is the method of leaf arrangement, so marvelous the adjustments that all plants must make to insure ample light, that it is not inaptly called leaf mosaic. As we shall see in the chapter on Plant Distribution, particularly in forests, certain variations or partial failures of the process have far-reaching results.
If leaves did not perform this most important function to perfection, all animals, including man as well, would perish, and it would almost seem that their obligation to us and the plant world might stop there as long as their success in reaching the light is so overwhelming. But there are no union hours of labor, no regulation as to the kind of work leaves may perform, and some actually reach out for new tasks to do, and do them. In one, our common pitcher plant, the leaf, as is implied by the name, is formed into a slender hollow pitcher, wide at the mouth, but narrow at the base. Inside the pitcher are slender downward-pointing hairs so arranged that an insect may crawl in, but never out. The lower down the luckless insect gets the more certain is its death, and, to clinch matters, there is a tiny pool at the bottom where it is not only drowned, but, due to the composition of the mixture in the pool, digested. Only a very few plants can do this; only a minute fraction of the world’s vegetation can digest animal matter. Some experiments on the pitcher plant, which grows in bogs, show that it will digest bits of beefsteak dropped into the liquid at the base of the pitcher.
In the East Indies and in Africa there is a pitcher plant—in fact, scores of varieties of them—which grows up on the branches of trees. In this case the pitcher may be as long as some of our American kinds, often twelve to eighteen inches, and many of them are attached to a slender leafstalk two to three feet long, by which they hang suspended. Insects, literally by the thousands, are caught in these gaudy traps, for many of the pitchers are beautifully colored, and near the opening they secrete a sweetish liquid that lures their prey. They are, in fact, such curious and handsome plants that they are commonly grown in greenhouse collections.
Nature sometimes finds still other ways of using strange and curious-shaped leaves, and in our American bogs is a group of plants, also insect digesters, still more unusual than the pitcher plants. In bright sunny places in open bogs one may often find small reddish, glistening plants, called sundews, usually only a few inches tall, covered with sticky hairs. In fact, the glistening is due to the secretion of the sticky substance, a tiny drop of which may be found at the end of each hair. Flying insects are caught in these leaves, and, as a fly on fly paper, the greater the struggle the more involved does the insect become among the sticky threads. Once caught by such a plant, escape is practically impossible.
Lying in ambush for chance insects, as these sundews and pitcher plants do, may seem nearly the limit of what is to be expected of leaves. Merely to be always on the job, with a plentiful supply of insect digester, might seem to be all that could well be expected from what, after all, are only modified leaves. But nature’s devices are infinite, and there are still other ways to accomplish the apparently impossible. In a small section of the southeastern States there grows a plant that not only lies passively in wait for insects, but actually captures them. This flycatcher, known as Venus’s flytrap, has two valves to the leaf blade, supported on a stout broad stalk so arranged that their fringed surfaces face each other. If an insect—and many do—alights between these valves, they close together rapidly and the prisoner is hopelessly caught by the interlocking marginal bristles that fringe each valve. In this case there are glands on the face of the valves, against which the live insect is tightly pressed, and which secrete a digestive fluid. When nothing remains the valves slowly open and are ready for the next victim. They may be made to close by slight irritation with a lead pencil, and it is the impact of the insect that releases one of the most curious examples of movement in leaves known to us. There are a few other plants in different parts of the world that by still other modifications of their leaves catch and digest insects, but none of them are to be considered as “insect eaters,” or other names implying that they have definite designs on the life of passing insects. The process is sufficiently remarkable, the success of the operation so sure, that there is nothing gained by attributing to such plants, as many have done in the past, malignant characters that are possibly confined only to man. The whole wonderful process is more reasonably explained by realizing that all these insectivorous plants are so by virtue of necessity, that many of them are bog plants, which are often hard put to it to get suitable food, and that the extraordinary change of shape and function is but one more contribution of leaves to the economy of nature.
In dry or desert regions, where the conservation of moisture is essential to plant growth, water storage by leaves is nearly as great an aid to the plant as we have seen it to be in the stems of cactus, South African spurges, etc. Our common century plant, whose leaves are, in some kinds, a hundred times thicker than in ordinary foliage leaves, is a good example of leaves adapted to water storage. In our southwestern deserts hundreds of species of plants can exist only by virtue of the fact that their leaves are so changed in their form or structure that they serve as reservoirs for water storage. This may be accomplished by thickening, or it is more often contrived by a thick coating of hairs. The surfaces of thousands of different kinds of leaves are clothed with hairs either on the upper or lower side, or sometimes on both sides. In many cases they are quite obviously protection from too rapid drying out of the leaf. In others, as in the nettle, the hairs secrete a stinging substance which seems to insure the plant against grazing animals.
Leaves, then, are for something more than to provide the beautiful foliage which is their most spectacular accomplishment. So varied is this in its beauty, from plain green leaves to the wonderful coloring found in begonias, coleus, and many other garden plants, that the sheer beauty of the panorama of foliage is likely to blind us to the more important uses of leaves. First of all must we consider them the factories, in which night and day are produced the food of all plants and most animals. Then in certain cases we have seen that, by every ingenious device known to nature, they perform other special work, such as helping the plant to climb where that is necessary, catching or even capturing insects and digesting them when that peculiar service is demanded of them, and, finally, serving as storage reservoirs in regions where water is scarce. Probably no part of the plant works so unceasingly each season at its varied tasks. In the autumn, dropping to the forest floor, its decomposition furnishes still other food for the plant, and, to crown all, this busy life and by no means unprofitable death leaves behind it, as a promise for the continuance of the work, a snugly protected leaf bud which will repeat the process the next season.
THE FLOWER
While the plant’s and, consequently, our debt to the leaf is seen to be tremendous, it cannot be ignored that, if plants produced nothing but leaves, the end of all plant life would come with the death from old age or disease of the present generation of plants. Except for those kinds that reproduce themselves by division or extension of their rootstocks, which bear buds, there would be no provision for increase. As only a comparatively small number of plants can reproduce by this method, it is obvious that something more must be provided to secure new generations of plants. Flowers, and the fruits and seeds which inevitably follow them, do this. All plants, with some exceptions to be noted later, produce flowers at some time in their life. In the case of the century plant, only once, after which they die. But except for ferns, mushrooms, seaweed, yeast, bacteria, and some other forms of so-called flowerless plants, a flower or blossom is to be found at some stage in the life of all plants.
If we examine the leaves of a goldenrod, we find that they are large below and diminish in size toward the top. Just below and among the flower clusters they are so much reduced in size and often changed in color that they cease to be ordinary foliage leaves, and are known as bracts. The occurrence of bracts is nearly universal in flowering plants, and they form not only an apparently transitional stage between leaves and flowers, but an actual one.
In a complete and perfect flower there are, at the bottom of it, a row of green leaflike sheaths which
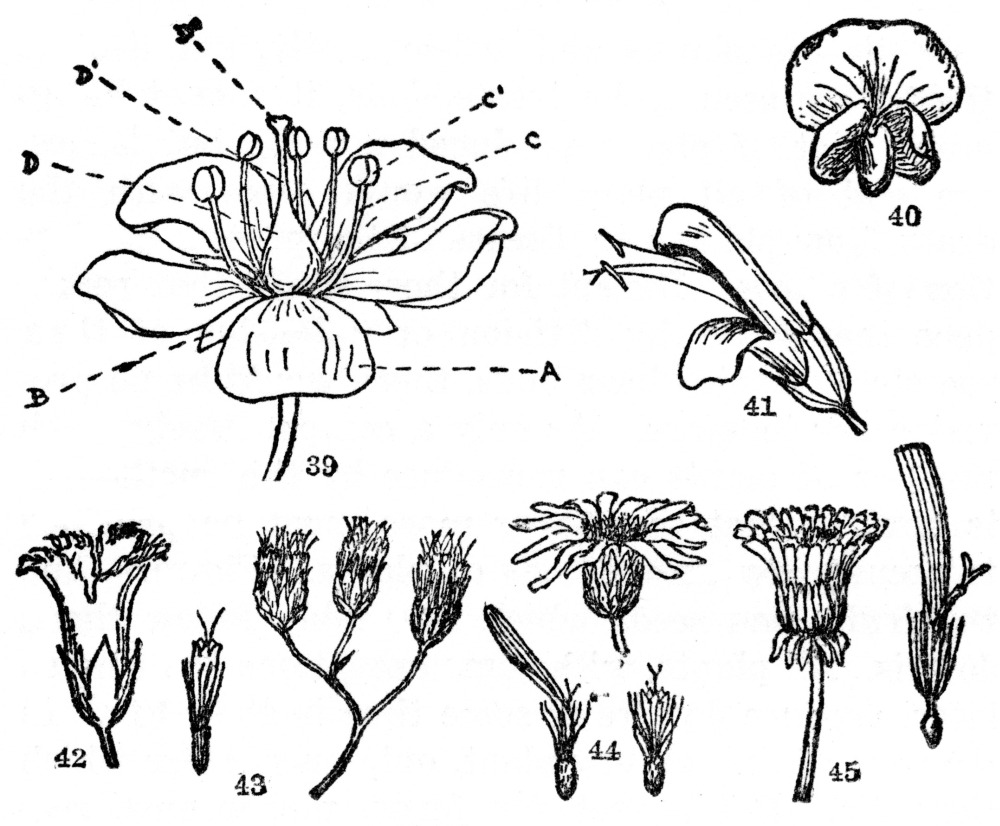 FIG. 39-45.—THE FLOWER
FIG. 39-45.—THE FLOWER
Fig. 39. A perfect and complete flower. A, petals, all of them forming the corolla; B, sepals, all of them forming the calyx; C, the stamen, composed of (C) the filament, and (C1) the anther, which produces the pollen; D, the pistil, consisting of the swollen base (D) the ovary, a slender shank (D1) the style, and the swollen or branched tip (D2) the stigma. (H. D. House, “Wild Flowers of New York.”) Fig. 40. Typical flower of the pea family. Two petals unite to form the keel (below), two more unite to form the wings (center), the remaining and larger petal forms the standard. In most plants of this family the stamens and pistils are concealed within the keel. Fig. 41. Two-lipped inequilateral flower, common in such plants as Salvia, Snapdragon, etc. Note the united calyx and corolla. Fig. 42. Gamopetalous or united and regular corolla of the Fringed Gentian. Figs. 43, 44, and 45, flowers of the Compositæ or daisy family. Many small flowers grouped in heads and usually surrounded by one or more series of bracts. Fig. 43. Flowers all tubular, the small one at the left being an individual flower. Common examples are Boneset and the common garden Ageratum. Fig. 44. Flowers both tubular and with rays, the tubular in the center and the rays on the margin. Below is an individual tubular flower on the right, and on the left an individual ray flower. Note that its five united divisions correspond to the five petals in other plants. Common examples are the daisy, sunflower, black-eyed Susan, etc. Fig. 45. Flowers all ray flowers, an individual one at the right. The Compositæ with only ray flowers usually have a milky juice and have often been grouped in a separate family, the Cichoriaceæ. Common examples are dandelion, chicory, and lettuce.
surround and often half inclose the brightly colored petals within. This outer covering of flowers is called calyx (Figure 39 B), the individual parts of it, where they are separated, sepals. Their chief use is to protect the interior petals while they are inclosed in the bud. The calyx may or may not have bracts just underneath it, as it does very conspicuously in the case of the flowering dogwood, whose white “flowers” are really only brightly colored bracts. The transition between bracts and calyx is not difficult to see in many plants, and where it is impossible the evidence from their internal structure confirms what our eye might be inclined to doubt.
Just inside the calyx is what most people call the “flower,” which is really composed of more highly colored sepals, but which we call petals (Figure 39A). Where these are joined together the collection, which forms tubular flowers like the lily of the valley, is called a corolla. It is, of course, the petals or corollas of flowering plants that give our landscapes their greatest beauty, their most gorgeous coloring. While this from one point of view amply justifies a prodigal nature in strewing the earth with beautiful flowers, the true value of the color to the plant is in quite other directions, which will be explained a little later.
Toward the base of the corolla, or sometimes on the petals or sepals, may be found a series of slender appendages, usually threadlike or a little thicker, crowned at the top by a distinctly large knob. The individual appendage is known as a stamen (Figure 39 C), its threadlike portion a filament (Figure 39 C), and the knoblike top an anther (Figure 39 C1).
Directly in the middle of the flower there is still another organ, usually swollen at the base, slender in the shank, and either thickened or branched at the tip. This central part of nearly all flowers is called collectively a pistil (Figure 39 D), its swollen base an ovary (Figure 39 D), the slender shank the style (Figure 39 D1), and the thickened or branched tip a stigma (Figure 39 D2). A perfect and complete flower, then, is composed as follows:
Calyx
or sepals, | + | Corolla or petals, | + | Stamens consisting of filaments and anthers, | + | Pistil consisting of ovary, style, and stigma. |
The stamen is the male organ of reproduction and the pistil the female. The actual process of fertilization, pregnancy, the forming of the fruit and later the seed, and the latter’s birth of a new plant, comprise one of the most fascinating of those provisions of nature which secure the perpetuation of the plant world. In the life history of even the commonest weed along the roadside there is this constant renewal of life by sexual reproduction, just as in animals and in man. In the chapter on “How Plants Produce Their Young” will be found some account of this supreme function of flowers, after which, as if their usefulness were over, they wither and perish.
Not all flowers are perfect or complete. Some lack petals, as the buckwheat, where the colored calyx replaces petals. Others have neither calyx nor corolla, as in the sycamore or plane tree. Most plants, however, have both calyx and corolla. In some very few plants certain of the flowers have no stamens, when they are said to be pistillate or female flowers, and certain others have no pistils, when they are called staminate or male flowers. In other words, the sexes are in different flowers in the same cluster or plant, as is true of the walnut and hickories, when they are said to be monœcious. In still others the sexes are on entirely different plants, in which case they are diœcious, as in practically all willows. In the latter case there are pistillate or female plants and staminate or male plants.
While it is a commonplace that peas do not look like daisies, nor a carnation like a rose, this simple observation does not begin to tell us of the wonderfully different flower shapes and colors that are to be found along any roadside. The perfect and complete flower that we have been studying is quite regular, composed as often as not of four or five petals, as many sepals, with five or ten stamens and perhaps a single pistil. Yet there is literally no limit to the variations from this scheme, and some of these must be understood here in order that the life-histories and behavior of plants discussed in later chapters may tell their full story.
The figures on page 44 show a regular flower, with five separate petals and sepals (Fig. 39). Such flowers are said to be polypetalous, i.e., separate petals. Sometimes three of the petals are larger, two smaller, in which case the flower is lopsided or, as it is said, inequilateral. Again all the petals are united to form a regular and equilateral tube, as in lily of the valley, when they are gamopetalous, i.e., united petals (Figure 42). As we shall see in the chapter on Plant Families, this is a distinction between two great groups of plants, as important in their classification as negro and white man are in classifying humans.
In peas, beans, the locust tree, and related plants the petals are much changed to form an irregular flower, with a keellike or prow-shaped part made from the uniting of two petals. Two more unite to form the wings, and the remaining and larger petal forms the standard. Figure 40 and the explanation under it illustrate this unusual form of flowers.
Our common garden salvia shows still another type of flower, which is tubular and irregular (Figure 41). There is an arching, hoodlike structure at the top overhanging a lower lip. This kind of irregularity is common in thousands of different sorts of plants and, usually, it is a device to insure fertilization of the flower by insect visitors. So necessary are these for pregnancy in many plants, that an orchid, once discovered in Madagascar with a tube eighteen inches deep, puzzled the botanists, who were unable to understand how the plant produced seed in the absence of any known insect with a tongue as long as that. Darwin said at once that such an insect would one day be discovered on that island. Years after, Baron von Humboldt, a German naturalist, found the insect and explained the mystery.
Perhaps there is no feature of plant life that shows such an amazing amount of variation as the forms of flowers, and while only a few of the simplest deviations from the normal have been discussed here, it must not be forgotten that this infinite variety is a reflection of the ingeniousness of nature in securing a plentiful supply of seed. Form, color, the secretion of sweetish nectar, the night or day blooming of different kinds of flowers, every device that will make fertilization certain, by the flower itself, by insects, or even by the wind, is used in such prodigal fashion, that we come to see the importance of it to all plants only by a realization of the complexity of it and the provisions against its failure.
One apparently most lavish method of securing fertilization is the arrangement of flowers in clusters. While many flowers are quite solitary, the great mass of individual plants produce a few or dozens, or even hundreds of flowers—in fact, certain relatives of the common carrot may produce over a thousand flowers in a single cluster. The form and plan of arrangement of these clusters follows a rather definite scheme, and here, as in the case of leaves and parts of individual flowers, the figures tell the story better than words. In the common dandelion and daisy, and their thousands of relatives, the “flower” (Figures 43-45), as commonly understood, is really composed of scores or even hundreds of true flowers in each head. In the case of the daisy the yellow center, if picked apart, is seen to be really made up of scores of tiny tubular flowers, each just as truly a flower as a single rose. The rays, or what are incorrectly called “petals,” which fringe the golden center with white, if carefully separated and examined closely, will be found to be also a complete flower, the true petals of which are all joined to make the strap-shaped ray. If one looks sharply, the united edges of these petals may be seen by the ridges or channels that represent their joined edges. Because plants of this sort produce two sets of flowers in each head, one conspicuous by its brightly colored rays and with another tubular set in the center which makes doubly certain the fertilization and seed supply, they are considered the most highly developed of all plants. It is not a close aristocracy, nor an exclusive one, for over eleven thousand different kinds of plants, scattered all over the world, have their flowers arranged in this fashion or some slight modification of it. They possess, above all others, the certainty that there will be no slip in their fertilization, pregnancy, and subsequent birth of a new generation. Because this is the great object of all flowers, and these daisylike plants have brought it to such perfection, they are most surely to be classed as the highest type upon the earth to-day.
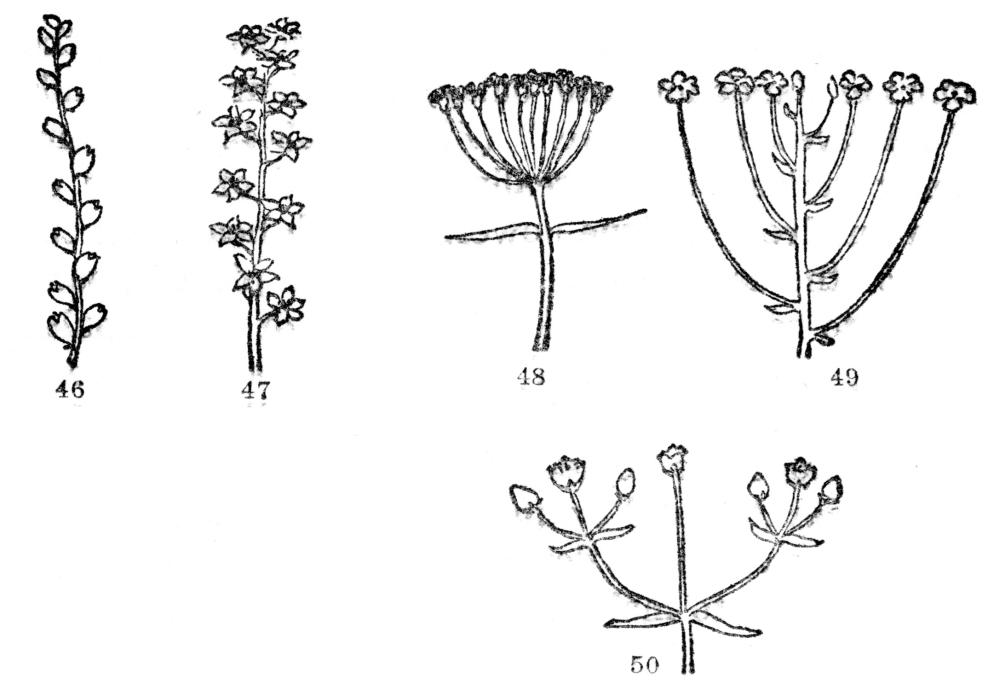 FIG. 46-50.—TYPES OF FLOWER CLUSTERS
FIG. 46-50.—TYPES OF FLOWER CLUSTERS
Fig. 46. A spike, the individual flowers attached directly to the common stalk. Fig. 47. A raceme, a spikelike cluster where individual flowers are stalked. Fig. 48. An umbel, the individual flower stalks all arising from one point. Fig. 49. Individual flower stalks of different lengths but the cluster usually flat-topped (corymb). Fig. 50. A flower cluster in which the end of the stem is terminated by a flower from the base of which side branchlets similarly tipped with flowers arise (cyme).
While highly irregular flowers are common in nature, conspicuous examples being the orchids in any florist’s window, or the milkweeds along the roadside, they can nearly always be seen to have various changes in the shape of their petals, or sepals, or stamens, or pistils, which are adaptations to their mode of life, but which always result in fertilization. Some plants, true monstrosities of nature, are not only far from having the usual arrangement of flower parts, but they even produce increased numbers of one part at the expense of others.
Double buttercups, and hundreds of our most beautiful garden blossoms, have been rescued by cultivation or the arts of the gardeners. Some roses seem to be practically all petals, but for every increase of petals there must be a decrease of some other part of the flower, and more often than enough it is the stamens and pistils that lose out in this transformation. Just as there is a decrease almost to the vanishing point in the birthrate when people become too effete and cultivated, so in plants there seems to be a point beyond which they cannot be pushed without suffering partial or often complete inability to produce young. The more highly they have been developed, oftentimes the greater their beauty, the less able are they to see to it that the chief function of flowers is accomplished. Such garden plants are increased by root division, cuttings and other arts of the gardener. Naturally true double flowers are almost unknown in wild plants, and the habit seems to have been brought about by too easy a time of it, too little struggle, too much food, or by any other of those things that produce effete but beautiful things, charming in their way, but of no significance in the sturdy struggle for existence that all wild plants must meet or perish. Another curious modification of a flower bud is cauliflower. Here the bud has been so developed, its calyx, sepals, etc., so transformed that the large, cabbagelike head, produced at the apex of the main stem of the plant, has by so much lost all semblance of a flower that it is actually a vegetable.
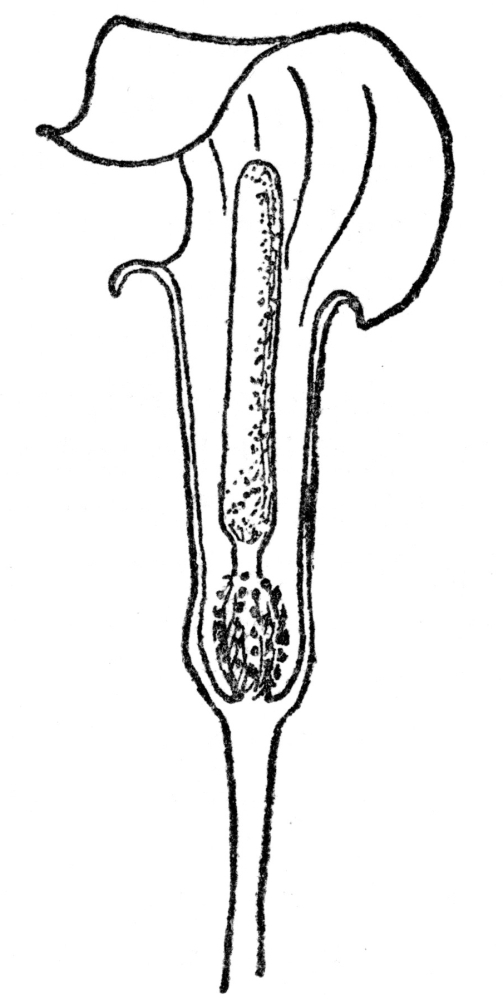 FIG. 51.—FLOWER ARRANGEMENT PECULIAR TO THE ARUM FAMILY
FIG. 51.—FLOWER ARRANGEMENT PECULIAR TO THE ARUM FAMILY
Fig. 51. The outer leaflike tubular or hooded spathe surrounds in our common Jack-in-the-Pulpit a clublike spadix, upon which are crowded the tiny flowers.
No feature of a landscape gives us more pleasure than its flowers, over which poets have sung and artists have painted their most charming pictures, even a musician has composed a very beautiful piano piece, “To a Water Lily.” But their true place in the scheme of nature has a deeper significance: the wonderful color and symmetry of their parts, the plan of their arrangement, their transformation into curious forms, like the Madagascar orchid, and hundreds of others—all these point to their supreme function, an act of self-sacrifice comparable only to the fall of a leaf when its task is done. Petals, too, wither and die when the fertilized ovary, already a mother, begins the slow process of maturing its young and the end of the flowering stage is reached. Such a climax is this in certain plants that the whole plant dies, as we have already noted in the case of the century plant. The toddy or wine palm of India, often sixty or seventy years old and more than a hundred feet tall, flowers only once, and, as if in recognition of the fact that it has done that for which it grew, slowly dies as the seed ripens. More humble annuals, like buckwheat, and hundreds of others, live only one brief growing season, produce flowers and seeds, and then die, leaving behind them the only means of perpetuating their kind. The dormant seed carries over the winter the life they were themselves unable to maintain, as perennials and woody plants do in their buds.
THE FRUIT
The number of different kinds of fruits that one can buy even in the greatest markets in the world is so small, compared to all fruits that are annually produced by plants, that they might almost be likened to an ear of corn as against a Missouri cornfield. If, as we have seen, all flowering plants must produce fruits, then what we commonly call such can be only a fraction of what actually makes up nature’s annual harvest. It follows that fruits often occur in unfamiliar disguises and, as we shall see presently, some of the things we have been calling fruits may be so only partly, if at all.
Disregarding what we call fruits and looking at it from the plant’s point of view, a fruit is anything in which, or upon which, a seed is developed or ripened quite without regard as to whether it is edible by man or not. As the ovary is the female organ of reproduction and contains the yet undeveloped seed, it follows also that fruits are practically always a development of some part or modification of the ovary or the upper end of the flower stalk upon which it rests and from which it is often scarcely separable.
Familiar enough is the distinction between dry fruits, such as a pea pod and fleshy ones like oranges, and this quality of being fleshy or dry is practically universal. Among fleshy fruits a few well-known types may be mentioned, such as the orange, tomato, grape, gooseberry, and cranberry, all true berries. There are, of course, thousands of less familiar examples of berries, but, whether with a hard rind as in the orange or not, they are a direct development, or often a mere swelling of the ovary, with sometimes the adhering calyx, and contain the seed. In apples and pears, known as pomes, the fleshy part is a development of part calyx and part the receptacle upon which the ovary is supported while still in the flower. The ovary in these fruits is the parchmentlike interior which contains the seed. Plums and cherries, which have a single stone, instead of numerous seeds buried in the flesh, are known as drupes. These familiar examples are matched by thousands of others of which we hear nothing, all drupes and all formed directly from the ripened ovary and without much change, except the increase of size, juiciness and large development of the tiny immature seed, now transformed into a stone. In the watermelon, pumpkin, and related plants, is still another kind of fleshy fruit, called a pepo. All of this, including the hard rind, is transformed ovary and calyx completely incorporated, and forming in the pumpkin perhaps the largest fleshy fruit known. In a considerable number of plants there is not a single ovary, but several, or in some cases many. These occasionally all develop into what is called an aggregate fruit, of which examples are the blackberry, mulberry, magnolia, and many others.
While it would be logical to think that these fleshy fruits were designed to make delicious food for man, that, in the light of what we have seen to be the real function of the flower, is an assumption which, while flattering, is far from the truth. It is much more certain that fleshy fruits help plants in the dispersal of their seeds and that this fleshy, juicy character is just one more device of nature to see to it that not only do plants produce seeds, but that the seeds are carried and so spread the plant over considerable areas. Birds and animals eat such fruits in enormous quantities and, in fact, bird migrations are thought to be not so much response to winter cold as to the fact that fruits are scarce then. When it is remembered that some birds make tremendous flights, often over 10,000 miles in a few days, their capacity to spread seeds through their droppings may be imagined. In the chapter on plant distribution some truly remarkable cases of such seed dispersal will be given.
The chance of having seed carried great distances, because it is embedded in a fleshy, often brightly colored fruit, would seem to put plants having dry fruits at a disadvantage. Birds and animals cannot be expected to look after the dispersal of those fruits that are neither tempting to the sight nor to the taste. And it must be confessed that quite other qualities in dry fruits insure their dispersal. Some are so nutritious, like the acorn, that thrifty squirrels store them over the winter, as they do many other seeds which are harvested from dry fruits. Various grains are often so stored by man, and rice, wheat, buckwheat, and other cereals are common cases. In nearly all grains the seed fills so completely the fruit that cereals are very generally, but mistakenly, called seeds. A grain of wheat or corn is just as complete a fruit as a watermelon. Only its outer coat and inner seed are so closely welded together as not to be usually recognized as a fruit, with the seed inside.
One of the commonest types of dry fruit is the capsule (Figure 53), well named, as it is almost an exact counterpart of the capsule of the druggist, in that it is in many cases composed of a lower part and an upper, usually merely a domed lid. Others again, instead of splitting around the sides, split from top to bottom. Still others, as peas and beans, known as legumes (Figure 57), are pods that not only split lengthwise, but have no central partition, as do many other fruits of the same general type. When the seed is ripe nearly all pods and legumes finally split open, and the seed or seeds tumble out. A few, as in the violet and touch-me-not or jewelweed, apparently realizing that merely to spill out ripe seeds at the proper time will not spread the species very far, open their fruits with a sudden explosion and literally shoot their seeds considerable distances. The artillery plant, commonly grown in greenhouses, a delicate feathery herb from tropical America, opens its flowers with a report like a toy popgun and shoots its small pollen grains for several feet, but not its seeds as stated by some.
But many fruits do not open at all and seem to be at the greatest disadvantage in the effort to insure
 FIG. 52-60.—TYPES OF DRY FRUITS
FIG. 52-60.—TYPES OF DRY FRUITS
Fig. 52. The strawberry. The fleshy part consists of the modified upper end of the flower stalk or receptacle, while the true fruits are the dry achenes on or embedded in the surface and popularly called the seeds. Fig. 53. A three-celled capsule splitting lengthwise as in the common Iris. Fig. 54. Fruit of the cocklebur, the hooked prickles of which are admirably adapted for clinging to the fur of animals. Fig. 55. Pods of a plant of the Mustard family, which split down both edges, unlike the true peas, which split down only one edge. Fig. 56. Two types of achenes of the daisy family tipped with plumed bristles, greatly aiding their carriage by the wind. Fig. 57. Common garden pea—a typical legume. Note that it splits only on one side. Fig. 58. The samara or two-winged fruit of the maple. Fig. 59. The samara or single-winged fruit of the ash. Fig. 60. The dry two-pronged and bristly fruit of the unicorn plant (Martynia), admirably adapted for dispersal by animals.
dispersal of their seeds. Greater food value to birds and animals overcomes this in some kinds, and another help is that some fruits of this sort are covered with hooked prickles or barbs (Figures 54 and 60). The common weedy burdock, the barbed fruits of which may often be found sticking to the fur of animals in great quantities, is a case in point. There are whole groups of plants that rely on this method for seed dispersal, notably the avens, tick-seed, tick trefoil, and many shrubs in the tropical regions.
Where the fruits are neither barbed nor very good to eat, and so apparently doomed to be more or less permanent stay-at-homes, nature has provided some of them with the proper equipment for flight through the air. Winged fruits like the maple are to be seen on any windy day during their season scurrying before the breeze, and consequently spreading their kind over considerable distances. In the maple there are two wings, joined at the base where the seeds are embedded in the wings, and the fruit is known as a samara (Figure 58), or key fruit, from a slight resemblance to an old-fashioned key. Ash trees bear fruits that are a slight modification of this type and may be carried considerable distances by the wind (Figure 59).
In the dandelion, daisy, and nearly all its thousands of relatives, this faculty of setting sail in the air has been carried to the greatest perfection, just as we saw its flowers were. In this family of plants, the largest in the world, the fruit is mostly tipped or surrounded by a small collection of very fine bristles. The fruit, known as an achene (Figure 56), is so light that with the added buoyancy of this tiny collection of down it can be transported great distances. Some have been known to fly hundreds of miles in severe storms, and, as we shall see in the chapter on plant distribution, these tiny plant balloons have played a conspicuous part in spreading their kind over the face of the earth. Cat-tails also, together with many other plants, have this faculty and make up by its possession for the lack of fleshy or otherwise desirable fruits that might be carried. All achenes are not winged, those which dot the surface of the strawberry being imbedded in the luscious flesh, which is not really fruit at all. Only the achenes on the strawberry are true fruits, the fleshy part being merely a development of the upper part of the flower stalk and not of the ovary (Figure 52).
Fruits, then, cannot be restricted to the common understanding of them. They are transformations of the ovary, in which or upon which seeds are nursed, and upon which most plants depend for the dispersal of their seeds. We shall see later on how fruits have fulfilled their destiny, how some are fit for their true function only when they have been eaten by birds, and when some digestive juice has released them from the impotence they would suffer without being eaten, how a whole forest has been changed in the West by the busy activity of squirrels upon the fruits and seeds of a single kind of fir tree; how the fruit of the coconut palm has been spread throughout the tropical world because it can float in the sea securely protected from injury from salt by the impervious coverings of its fruits.
THE SEED
As the final stage in the development of all plants is their seed, with the dropping of which they bid good-by to their fellows, it is not perhaps remarkable that in the seed of all flowering plants is the germ for the new generation. To seeds which may be as small as the mustard, so often mentioned in the Bible, or as large as the coco de mer, or double coconut, from the Seychelles Islands, often fifty pounds or over, is intrusted by cunning nature the one final and most important act in the whole kingdom of the plant world. Nearly all plants would die off forever if seeds did not have in them the germ of life, apparently quite dead, but actually only dormant. This living germ may persist for years, sometimes even a hundred years, and yet with the proper conditions it never fails to sprout.
Seeds have inside them a tiny plantlet folded and ready to grow when the seed splits to release it. Also, in the seed is stored up food to sustain the new plant until such time as its own roots begin to act. This young plantlet is known as the embryo, and to this all actions of the seed are subservient.
As the seed splits, and the young plant develops its first leaves and rootlets, there is shown one of the most remarkably uniform tendencies in plant life. In all plants with net-veined leaves the young plantlet starts life with two leaves, or cotyledons, as these first leaves are called, and this whole group of plants are thus known as dicotyledons. In plants with parallel-veined leaves the young plantlets start out with a single cotyledon and are therefore called monocotyledons. In only the pines, spruce, and a few other evergreen trees the seedling plants have several cotyledons and are known as polycotyledons. All the flowering plants in the world belong to one of these groups, so that merely to see the germinating seed tells the story at once. The linking of parallel-veined leaves and a single seed leaf, and net-veined leaves with two seed leaves, is also associated with very definite arrangement of their flower parts, their method of growth and other characters. Something has already been said of this in the discussion of stems and leaves, and more will be found in the chapter on plant families. No more beautiful example of the plan or scheme of nature is to be found than these characteristics of all plants, and in seeds we find the first hint as to which army the plant will join, under which banner it will fight, and under what generalship it will develop. Nothing tells us so much as these first seed leaves, pushing their way up through the soil and revealing, as they burst above ground, to what place in nature their destiny will consign them.
Flowering plants, which make up the bulk of the vegetation of the earth, have been discussed in some detail, not only because they furnish us with all the things that make life possible, but also because they show perhaps better than anything else the division of labor, all striving for one end. Roots, the food gatherers. Stems, the framework for the foliage and its means of reaching the light, or as a storage house for reserve food. Leaves of many kinds, all factories working night and day to make the necessary food. Flowers of every hue and shape to lure insects, or by other means secure union of male and female. Fruits to ripen the result of this mating of the sexes. And, finally, the seed carrying with it the yet unborn life. Each part occasionally losing itself in order that the end may be accomplished, many of them changing their form or even their function where that is of advantage, all in their separate ways doing their task, the end of which they cannot see, and the fruits of which they will never enjoy. Nowhere is it so true as in plants that to save oneself there must be the capacity to give oneself. Untold millions of leaves fall, or trees crash down, or seeds are developed, each fulfilling their destiny which is to insure the perpetuation of their kind. As we shall see later on, there are many mistakes, many apparently futile attempts, thousands are wiped out that one may be saved, and in the past multitudes have gone out forever. Yet the result of it all is the plant world as we know it to-day, each kind struggling to increase its sphere of influence, or to cover more of the earth’s area. The combat between different kinds is inexorable, yet the capacity for sacrifice on the part of different organs, in order that a certain individual kind may win, is literally beyond belief.
2. Flowerless Plants
In the light of what has been said about flowers it may well be questioned how anything can be a plant and still have no flower. The fact is that flowers as we commonly understand them are unknown in the plants about to be discussed, but that what corresponds to a flower, and performs the function of a flower all plants must and do have. In the case of most flowering plants the possession of flowers is one of the beauties of nature in its most resplendent mood, while in the so-called flowerless plants the functions of flowers are performed by tiny microscopic organs, even the existence of which has been only recently discovered. Because flowering plants produce their sexual organs in such a gorgeous setting, for all the world to see their matings they have been called phanerogams, which means literally visible marriage, while the flowerless plants which perform similar functions in more secret ways are called cryptogams, meaning hidden marriage.
These cryptogams or flowerless plants occur in far greater numbers in the world than flowering plants, but their size in most cases is very much less. Many individuals are so small, as in the case of bacteria, that a single one can only be seen after it has been magnified many hundreds of times by the microscope. Of the cryptogams some of the largest, certainly the most beautiful, and probably the best known are
THE FERNS
In nearly all woods one may find delicate feathery plants with graceful, usually much divided leaves that nearly always start up from the ground like a slowly opening, but somewhat fuzzy coil. (Figure 62.) Ferns, at least most of those that grow in America, uncoil their leaves in this way, almost without exception. The accompanying figure shows the procedure, and in addition to this character one may hunt in vain for flowers.
While they bear no flowers we already know that nature could not leave them with no means of reproduction without abandoning them to a childless old age and the consequent extinction of the race of ferns. So far from the truth is this that ferns make up a goodly proportion of the world’s vegetation, and there are many hundreds of different kinds known. The lack of flowers, of course, explains why ferns do not bear seeds which are matured in a fruit or ripened ovary.
On the back of the leaves of most ferns, along or near the edges of the finer subdivisions, one may
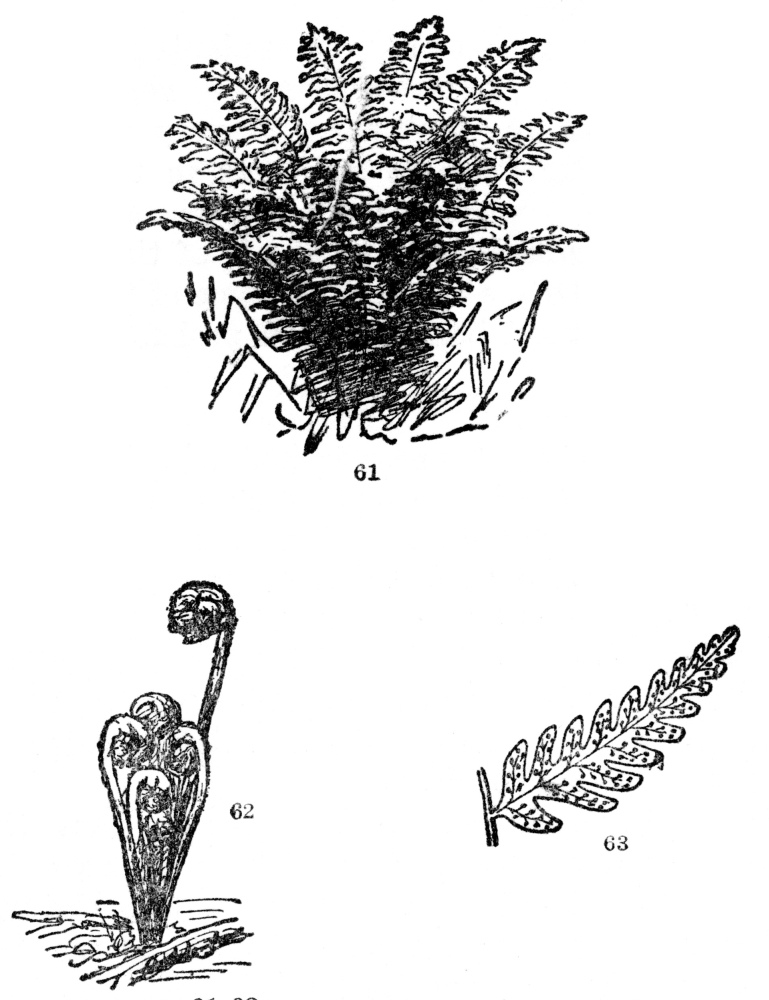 FIG. 61-63.—COMMON WOODLAND FERN
FIG. 61-63.—COMMON WOODLAND FERN
Fig. 61. A general view. Fig. 62. Its uncoiling spring condition. Fig. 63. The back of one of the smaller divisions of the leaf showing the collection of spore cases (sori). These are sometimes borne on special leaves, but in most of our American kinds on the backs of ordinary foliage leaves.
find, at the proper season, collections or rows of tiny, usually brownish dots. These contain often thousands of microscopic objects known generally as spores, and from this fact the dots are called spore-cases, or more technically sori. (Figure 63.) The process by which new plants are formed is a

Aerial Roots of Fig Trees Hanging Over the Edge of a Cave in the Rain Forest, San Lorenzo, Santo Domingo. (Photo by the author. Courtesy of Brooklyn Botanic Garden.)

Venus’s Flytrap, an Insectivorous Plant of the Southeastern United States. The fringed valves of its leaves close together when an insect alights between them. (Courtesy of Brooklyn Botanic Garden.)
somewhat complicated one, but the spores in these brown dots are the agency which makes reproduction possible, and the actual mechanism of it, one of the most interesting achievements in plant life, will be described in the chapter on “How Plants Produce Their Young.” Sometimes the spores are not borne on the backs of ordinary foliage leaves but on special leaves that bear, very often, nothing else.
Ferns are much like ordinary flowering plants; except for their lack of flowers, they have all the root, stem, and leaf characters of their more showy neighbors. While most of them have compound leaves, even sometimes twice or thrice compounded, a few have simple, narrow leaves without teeth, and one kind in tropical America has threadlike leaves. In many tropical rain forests, so called from their dripping wet condition, ferns form large trees, and these tree ferns are among the most graceful and feathery of all plants. There are, too, a few climbing kinds—one, called the climbing fern, is a native of the eastern United States. Then there is the walking fern, that seems to upset the statement that plants do not move as animals do. It sends out delicate runners that, rooting at the tips, form new plants, often several feet from the parent plant.
The characteristic of having, even in the simplest form, stems, leaves, and roots, with all that this implies in their internal structure, marks them off at once from all other flowerless plants. In ferns there is always some internal equipment for carrying food from one part of the plant, the roots, to another, and this ability is possessed by virtue of ducts or vessels through the stem and leaves. This system, found in all flowering plants and ferns, but nowhere else in the plant world, is called the vascular system, or literally, a vessel system. We shall see how important was the acquirement of this system of vessels, when we get to the chapter on the History of the Plant Kingdom. Its appearance upon the earth marks as important a stage in the development of plants as the dawn of a definite backbone did upon animal life.
Ferns, then, are vascular cryptogams because they do have conducting vessels in their stems, and they produce their young by a process of hidden marriage which will be described later. All other cryptogams or flowerless plants are without this system of vessels and are called therefore non-vascular cryptogams. Numerically they are tremendously important; upon them depend many manufacturing processes like bread making, brewing, and all arts using fermentation. But they are hardly recognized as plants by the general reader, and because of their size and the necessity of studying them with a microscope in order to understand their structure they will be treated here only briefly.
OTHER FLOWERLESS PLANTS
The remaining flowerless plants, having no duct system in their make-up, are, as we know, called non-vascular cryptogams. This is a general term for a very large group of plants, some quite obvious and well known like a mushroom, for instance; others so small or of such uncertain structure that they are not even well known by experts. This great mass of plant life, more numerous than all the other kinds of plants combined, contains many different forms, some of which are of gigantic size. A single plant of a certain Pacific Coast seaweed regularly exceeds in length the height of the tallest
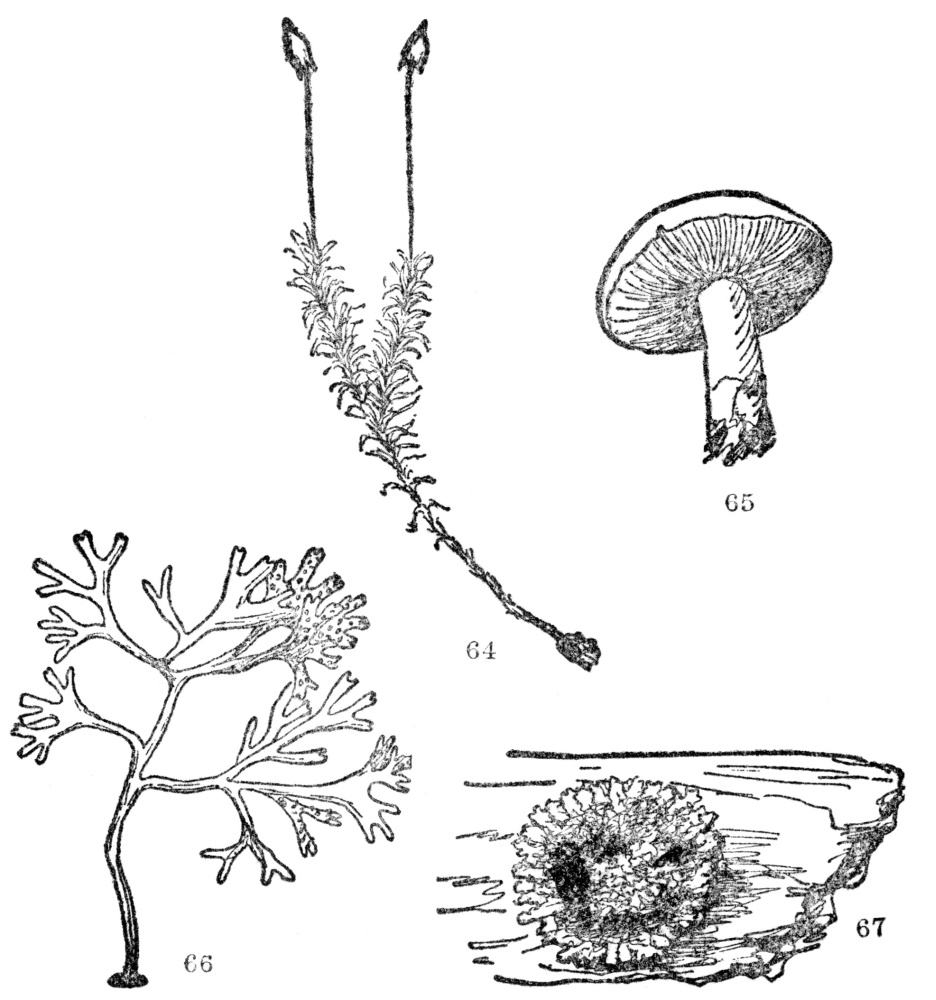 FIG. 64-67.—TYPES OF FLOWERLESS PLANTS
FIG. 64-67.—TYPES OF FLOWERLESS PLANTS
Fig. 64. A moss plant. Fig. 65. A mushroom, a common type of the fungi, which include also puffballs, molds, and many disease-causing microscopic organisms. Fig. 66. A common seaweed, a representative of the algæ, which include the green scum on the top of ponds, and the kelp from which fertilizer is now being made. Fig. 67. A lichen, a common cryptogamous plant on logs and rocks. Our native kinds are usually grayish-green in color.
known trees. And yet other inhabitants of the water, certain kinds that float freely, are microscopic in size. The latter occur in such enormous numbers that their tiny decomposed skeletons after dropping to the bottom of the sea form the diatomaceous earth, so much used in polishing machinery. The commercial product now comes from deposits of these skeletons laid down in past ages, which, due to changes in the land and water surfaces of the earth, are now found in Virginia, Nevada, California, and in Bohemia. All these must have been in the bed of waters long since gone, which teemed with these microscopic organisms. To-day there are over ten thousand different kinds known, yet so small are they that their dimensions are measured in thousandths of an inch!
Somewhat lower in the scale of life—and by this we mean simpler in structure—than the ferns are the mosses. (Figure 64.) There are thousands of different kinds, but everyone is familiar with the collective growth of the commoner sorts which makes the velvety mossy carpet in our woods. The individual plants are small, but in many kinds sufficiently large to be seen without a microscope. Most important of all, practically every one of them has the ordinary green color of the better known plants, and as we shall see in the section devoted to “Leaves as Factories for the Making of Food,” that stamps them at once as plants, if other things did not.
Mosses are almost infinite in their habits, some growing on the dry rocks or trunks of trees, many growing in moist woods, some in the water, and immense quantities of certain kinds in bogs. The peculiar bog mosses, known as sphagnum, play an important part in forming peat and perhaps coal. While mosses are otherwise not of much commercial importance, they are among nature’s most beautiful ground covers, carpeting many a nook and dell with a soft, velvety, almost cushionlike growth.
Although they are rather small, they appear to have a somewhat definite stem and tiny leaflike appendages of it, without, however, having the vascular system found in all ferns. Mosses might almost be considered miniature ferns, of which they are perhaps only simple ancestors. Their vegetative or green parts vary much in shape, size, and the arrangement of the tiny leaflike appendages, and while most of them are a beautiful bright green, nearly all the bog or sphagnum mosses are rather ashy gray in color. In most of the typical mosses there arises from among the vegetative growth of them a slender stalk, at the top of which is a small capsulelike organ. This contains the spores, and it is upon this long slender stalk and its spore-filled capsule, really marvelous in its internal structure and mechanism for the discharge of the spores, that mosses depend for their reproduction. As in the case of the ferns this process will be considered later, along with that of some other plants. This whole story of how plants produce their young, perhaps the most fascinating of any part of the study of plant life, is so fundamentally a part of their history and shows nature in her most maternal moods, that a special chapter will be devoted to it. There we shall see, as a whole, how these vastly different acts of fertilization and reproduction are, in different groups of plants, all responses to that insistent command for life, more life, in a never-ending stream.
The chief characters to remember about mosses are that they are very simple, but practically always green plants that have some differentiation into stem and leaf; that, while they have no vascular system, their structure and particularly the mode of reproduction suggests that they are not very distant from the ferns, and quite likely simple ancestors of them. These characters are of more importance than appears on the surface, as we shall presently see, for they mark mosses off from many other nonvascular flowerless plants which have quite different structure and altogether different mode of life.
If you will turn to the chapter on Plant Behavior and read particularly the sections on “Leaves as Factories for the Manufacture of Food” and “Borrowing from the Living and Robbing from the Dead,” you will see in the food habits of the plants there noted the great difference that exists between plants, like mosses and ferns, that have green coloring matter in them, and those we are about to mention that never do. The lack of this green coloring substance tells us at once that plants of this sort live only on the dead remains of other plants. In the case of these nonvascular flowerless plants there are certain modes of growth that, in some forms at least, are always associated with this scavenger-like food habit.
The common mushroom (Figure 65) is the best known of that large group of plants, called generally fungi, which produce no green coloring matter, have no leaves attached to a stem, and always live on decayed vegetable, or sometimes inhabit living animals, even man himself. The mushroom with its brownish stalk and buttonlike dome is familiar enough, but there are literally thousands of different kinds, a common sort forming “brackets” on the trunks of trees. While perhaps everyone would recognize these as plants, peculiar as they are in their often weird shapes and unusual as they nearly always are in their color, there are many minute kinds of fungi that scarcely anyone would even think of as a plant, and yet for better or worse they are incomparably the most powerful plants in the world. For upon these microscopic fungi man depends for many things. It is certain kinds of them that make the manufacture of cheese possible. They turn milk sour (pasteurizing milk is merely stopping their work), give to yeast its power of “raising” bread, all brewing depends upon them, every process of fermenting the juice of fruits for wine making or for whatever else, the decay of wood—all these processes and scores of others, whether for the good or evil of mankind, depend upon the work of these plants, any one of which is so small that a single individual must be magnified hundreds of times to detect it. Many of them are the “germs”—better called bacteria—that cause diseases like tuberculosis, cholera, typhoid, anthrax, and diphtheria. All surgeons wage incessant warfare against a host of them that attack wounds and form pus. They live in our intestines and have much to do with digestion, and unhappily with indigestion, so that we may be said to carry about with us a whole flora of them! Nearly all the diseases of plants, like the blight of potato and the rust on wheat, are caused by them. Some other kinds live in the soil, and many flowering plants depend absolutely for getting their food upon the work of these fungi. Unfortunately their minute size and consequently obscure mode of life demand technical skill and the use of the microscope to detect them, so we must leave them here, always keeping in mind that these smallest of all plants are charged with a power for good or evil; so far as man’s life is concerned, greater perhaps than all other plants.
While most fungi, particularly those familiar ones like mushrooms and puffballs, are inhabitants of the land, the remaining group of nonvascular flowerless plants are nearly all water plants. Most of the better known ones live in the sea, and as the wrack or tangle washed up on the shore we recognize them as seaweed. The algæ (Figure 66), which is a general name for such plants—and they live in the sea, in fresh water, and even on dry land—are, so far as structure is concerned, the simplest of all plants.
Those that are fastened to rocks are often beautifully colored, much branched, and many kinds bear small bladders that act as buoys. These coast seaweeds are generally of different colors, those nearest the surface being generally greenish, the deeper water kinds reddish or brown. None of these seaweeds are found at great depths, because the really deep parts of the ocean are almost, if not quite, dark. Seaweeds, and in fact all the algæ, have green coloring matter in them, even where this is masked by reds and browns, as is the case in some particularly showy kinds. As you will find in the section on “Leaves as Factories for the Making of Food,” no plant with green coloring matter can live in the dark. That is why seaweeds are not found in the great deeps of the sea, some of which are several miles below the shore line along the coasts, and are so cold and dark that neither plants nor animals can grow in them.
Those seaweeds that grow along the coast, and are uncovered by the retreating tides, are well known by everyone, but by far the greater number of algæ float without anchorage of any kind. One kind that has been torn from its anchorage occurs in such enormous quantities that off the coast of America it has formed literally a floating island composed entirely of dense mats of a species of seaweed. This place, known as the “Sargasso Sea” from the name of the seaweed forming it, was the terror of old mariners and Columbus’s ship was fouled in it for two weeks. The area occupied by the weed is several hundred miles long and wide, and while old sea yarns about ships being caught in it and never escaping are gross exaggerations, it is certainly one of the most curious of plant growths, due entirely to a nonvascular cryptogam.
Of those kinds that are never anchored the number is legion, and in addition to those forming the diatomaceous earth, already mentioned, there are many more. They form almost the only food of hosts of creatures of the sea, but because of their floating freely in the water, the consequent difficulty of collecting them, and their unusually minute size, little is likely to be known of them, except by the experts.
Other algæ are always found in fresh water and form the scum found on stagnant pools. Individuals of any of these are so minute that, while under the microscope they are of the greatest beauty, their structure must remain for most of us a sealed book.
Summary Of What Plants Are
We have now traced, in only the briefest fashion, the outlines of what plants are, reversing the order of nature in beginning with those most complex but best known, the flowering plants. As we shall see later, these are the climax of prodigal nature and are to be considered the end rather than the beginning of plant life on the earth. Then, and still more briefly, have we stopped to see those less known plants that produce no flowers, such as the ferns, mosses, fungi, and finally the seaweeds or algæ. These are all to be considered as the ancestors of flowering plants, the ferns the nearest to them and the algæ probably the most distant relatives. The development of plants from the minutest alga up to our most gorgeous flowering plant, is an infinitely slow and painful process. With many mistakes, with its pathway strewn with the wreckage of forlorn hopes and false starts, it is incomparably the most dramatic story in the plant world. Some of its details will be told in the chapter on the “History of the Plant Kingdom.”
Nor can we leave the discussion of what plants are without some mention of the thing that really makes up their structure, whether it be a microscopic bacterial organism or the Big Tree of California. For the unit of all animal and plant life is the cell. In its simplest form it is merely a minute sac with a definite wall and inside the wall is a substance known as protoplasm, literally protos, first, and plasma, thing formed. It is protoplasm that forms the living tissue of all plants and animals; it is life itself. No one has ever succeeded in making any, notwithstanding that many learned men have tried for years. Its inclosure in the cell wall, its power of self-division and consequent multiplication of the units, make up those first things about which most of us can never know much, but the end of which we recognize in the beauty of plant life all about us. For only under the highest powers of the microscope may cells be seen and studied. Just as bankers reckon mills as a definite unit of a cent, and yet none of them has ever seen a mill, so we must think of cells as the definite unit of all living things, although most of us will never see a cell. But, unlike the mill, cells may be seen by those equipped to see them, and this study, the development and grouping of them to form all the varied objects that inhabit the plant world, is known as histology. It is literally the internal history of plants and animals, and lies quite outside the scope of this book. What we must never forget is that whatever knowledge we have gained, either from the foregoing account of what plants are, or from our observation of them, is, after all, only a partial notion of them, as unsatisfactory as our estimate of what people really are, from merely looking at the outside of the houses in which they live. The outer form we may know and admire, the inner substance must ever remain for most of us a secret treasure house the value of which is certain, but the key to which we do not possess.