CHAPTER VIII: DISTRIBUTION OF PLANTS
WE have seen in the previous chapters how many and how varied are the activities of the plant world and in this final one we shall get a glimpse of what these activities have produced. All the delicate mechanism of food getting and the manufacture of starch; the fertilization of flowers by insects, the wind or water; the response to changing light and to climate—these and scores of other activities of plants have resulted in the present vegetation of the world being what it is. Here we see the final reflection or register of not one but all the kaleidoscopic evidences of plant response and activities and history working in harmony, or, as we shall see presently, sometimes in violent conflict, and leaving as the result the wonderfully varied vegetation that now covers the earth. If we could read aright the story of which the vegetation of any particular country is the silent narrator, it would tell us not only what happened in the past but what is likely to happen in the future.
Plants, by what amounts to a kind of fatality, are rooted to the spot where they grow so that, unlike animals, their rapid distribution appears to be almost impossible, and yet the tremendous distances that some species have traveled seem like a pretty successful protest against the fact of the anchorage of individuals to the point of their origin. It is more than a successful protest, for it amounts in many species to an active campaign for dominance, to the exclusion or extinction of less aggressive neighbors, so that in any field or meadow or forest there are silent struggles constantly going on. Some of these are so inexorable in their results that they change not only the frequency of occurrence of the individuals involved, but sometimes the whole type of vegetation.
The competition to occupy just as much of the favorable plant sites as possible has been much aided by many species possessing means for the dispersal of their seeds or fruits that are ingenious in the extreme. Some of these are written plainly enough in the structure of the seed and its wonderful adaptability for the peculiar conditions to which it will be subjected. Before considering some of these structures we may profitably see how some plants look after the dispersal of their seeds within their own limited sphere of action.
In hundreds of plants the ripened pods, instead of being erect as their flowers have been, are pointed downward about the time the seeds are ready to be released, and their harvest is sown, sometimes by deliberate movements, in the immediate vicinity. No great areas are captured by such plants, except by the slow process of successive generations extending their range a few inches or at most a few feet a year. The great bulk of all seeds never do grow into new plants, but in those that only shed their seeds close to the parent plant the opportunity to reach new sites is by that much restricted. The chance of the species getting very far afield except by slow invasion of the neighboring region is limited. A few of such plants show remarkable ingenuity in reaching the utmost distance possible, perhaps the most effective cases being those that shoot their seeds by explosive bursting of their pods. Nearly all the violets do this, often shooting seeds several feet from the parent plant. Many plants of the pea family have pods that are twisted, which upon splitting release the previously pinched seeds so suddenly that they are shot considerable distances. In the common witch-hazel (Figure 111), the seed is shot through the air often as much as thirty feet.
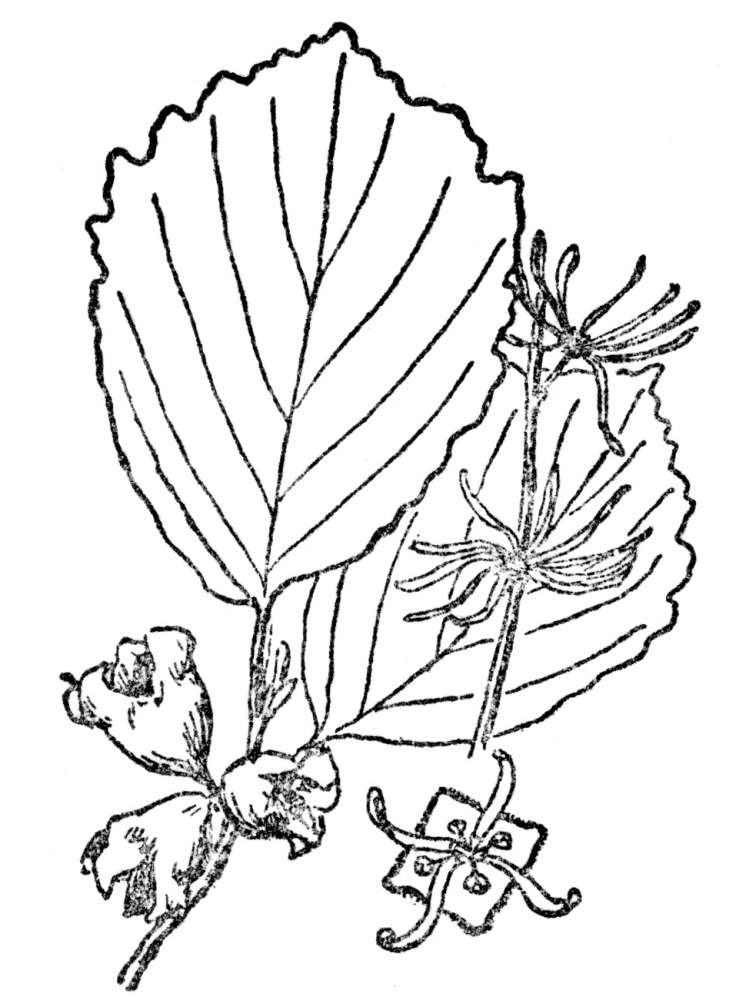 FIG. 111.—THE WITCH-HAZEL (Hamamelis virginiana) Is a fall or winter flowering plant which shoots its seed sometimes as much as thirty feet. Native of eastern North America.
FIG. 111.—THE WITCH-HAZEL (Hamamelis virginiana) Is a fall or winter flowering plant which shoots its seed sometimes as much as thirty feet. Native of eastern North America.
But with even the greatest ingenuity and the most explosive bursting of pods, most plants could never capture much new ground, and their very existence as a species is often contingent on their ability to spread, if these various methods by which plants shed their seeds were not aided by outside help. Some of these have had conspicuous and almost startling results.
1. Dispersal By Animals, the Wind, and Water
The seeds or fruits of those plants that are used for animal food are often carried considerable distances, while the thrifty squirrels’ burying of acorns is everyday knowledge to those who have seen them busily engaged in the making of winter stores, or the planting of new trees, often many rods from the parent tree. In years of plentiful seed production squirrels have been known to plant great quantities of seeds of the Douglas fir, thereby hastening the establishment of one of the greatest evergreen forests in the west.
While some seeds are destroyed by passing through the digestive tract of animals many are not harmed in the least. From over two hundred and fifty different kinds of plant seeds fed to a variety of birds over 80 per cent germinated perfectly after passing through their digestive tracts. And perhaps the most remarkable case is the seed of a pondweed said to be incapable of germination until it has passed through a bird. This plant grows in fresh water ponds in great quantities and is much eaten by wild ducks. From the stomach of one bird over three hundred such seeds were recovered. In the Eastern States a common feature of our farms is the red cedar or juniper scattered along fence rows, nearly all of which are due to birds roosting on fence rails and dropping the seeds after passage through their digestive tract.
We get some idea of the part birds play in plant dispersal when we realize the enormous number of them that make their flights twice a year, often over great distances. Wild ducks, in untold millions, travel from the far north to the tropics, each carrying their freight of seeds, sometimes as food and often mechanically clinging to their feet. The writer once saw at Gardiner’s Island and Montauk, Long Island, hundreds of thousands of tree swallows which feed on the fruits of the bayberry (Myrica carolinensis). So dense was the flock that they covered nearly every inch of the bayberry patches, and after eating no one can calculate how many million seeds they started off toward their winter home. Stopping as they do each day on their long flight southward, is it any wonder that the bayberry is one of the commonest bushes along the Atlantic coast?
With flights of birds stretching from the Arctic to the Antarctic, sometimes a single species making such a flight during its migration period, and hundreds of species making shorter flights twice a season, it is easily seen how birds can carry seeds for long distances. That they do so carry them is common knowledge and in eleven wild ducks examined by H. B. Guppy, he found nearly 300 seeds of bur reed, forty-one of pondweed, 270 sedges and 222 seeds that he could not identify. Nearly all these seeds germinated when sown, some sprouting more quickly than if they had not passed through the bird’s stomach. Some few seeds which would usually be dormant for one, two or even three years, have their germination unquestionably hastened by passage through birds, who may be looked on in some cases at least as “flying germinators.”
Some sea-flying birds have been captured over five hundred miles from land and seeds of a buttercup and of the sea blite recovered from their stomachs. One student of the flora of Spitzbergen has stated that nearly all the plants of that cold region have come from the northern part of the Scandinavian peninsula through their carriage by birds. One of the commonest plants of that region is the crowberry, which is a favorite food of birds. The plant is found throughout the Arctic regions and on high mountain tops to the south of it, such as the Alps, our own Adirondacks and White Mountains, and many others. There are some islands in the Pacific known to have received a few of the plants now growing on them by birds carrying the seeds from other regions, often thousands of miles away. Several species of birds are known to make sustained oversea flights from Labrador to South America, and one from Alaska to Hawaii. In such flights stopping is impossible, so that the carrying of fresh seeds and spores is always likely and probably more quickly accomplished than by any other means of transport. In New Zealand a northern European bird was once found with the seeds of two species of marsh arrow grass in its stomach, both of which germinated. The plants are native in the cooler parts of the north temperate zone.
Not far from New York, at Montauk Point, Long Island, there was found a few years ago some plants of the cloudberry, a kind of blackberry with amber-colored fruits, which is otherwise unknown in that region but is common enough in the Arctic and on mountain tops northward. That point of land, extending out to sea, is a favorite stopping place for migrating birds and to them was undoubtedly due the introduction of the plant so far south of its true home. Scores of similar cases could be cited which confirm the observations of naturalists all over the world that birds are among the greatest aids to plants in securing wide dispersal. Of course the factors of favorable or unfavorable conditions, once the seed is deposited in the new home, operate to keep plants from the tropics from settling permanently in colder regions and vice versa, but we cannot escape the conclusion that the sky is filled during certain seasons with millions of seed carriers that have, in many cases, populated their stopping places with foreign plants.
Other animals than birds carry fruits and seeds, besides those that eat them. Some fruits like the Martynia (Figure 60), are so arranged that no animal with fur can avoid catching some if they come in contact. The prickly fruits of cocklebur resulted in one species of that weed from the steppes of Russia being carried all over southern Europe in a comparatively few years. Tickseeds, Bidens, and all the hosts of plants that have prickles, burs, spines or what not attached to their fruits, have, by the possession of such devices, a better chance for dispersal of their fruits than those not so provided. Others, again, have various coatings of mucilage which stick to animals and thus help plants to overcome their chief drawback to dispersal—their anchorage at the place of birth.
THE WIND
The great trade winds, the violent hurricanes and monsoons of Asia, are all active and constant aids to plant dispersal. The fruits of many plants are provided with various devices to insure buoyancy in the air, such as maple, ash, and most seeds of pines and their relatives, birches, poplars, and willows, all of which may be carried distances of a few hundred feet in ordinary winds. In the buttonball tree, the milkweeds, fireweed, and many other plants there are feathery attachments or plumes that insure their seeds or fruits being carried very long distances indeed in regular or violent winds. But in the daisy family or Compositæ we find nearly all the eleven thousand species provided with a plumelike attachment of their light fruits, familiar enough in the dandelion, which may explain their being more widely distributed than any other plants. Some of them, however, are carried by animals, as they produce, in certain genera, barbed or hooked achenes.
The whole seed of some plants is so small that it can be lifted bodily by the wind, for instance Rhododendrons and many other heaths, one of the hanging pitcher plants of tropical Asia, many orchid seeds, and of hundreds of other plants. Many of these are less than one ten-thousandth of a gram in weight and, with dust and the spores of nearly all cryptogams, may be transported thousands of miles. When it is recalled that the dust from a volcanic eruption at Krakatoa in the Pacific was picked up on London window sills, and that some seeds and nearly all spores are as light as most dust, the wind as a plant dispersal agent becomes significant. In the western part of the United States over 800 million tons of dust are carried over 1,400 inches in a single year, according to estimates by J. W. Evans in his article on “The Wearing Down of Rocks.” Some effects of wind dispersal of seeds and spores furnish interesting data to the plant geographer.
In the Bahamas the natives speak of “hurricane grass” as a plant that was unknown on Great Bahama Island before August 13, 1890, when there was a great hurricane. Soon after this the sedge, which had been blown over from another island, began to be common on Great Bahama, where it is now thoroughly established. On Krakatoa near Java a violent eruption in 1883 completely destroyed the vegetation, covering the island with volcanic material. In thirteen years over sixty species of plants had arrived on the island, of which about twenty had been blown there as seeds or spores. Birds had carried about 7 per cent and the remainder had come by other means. No plants with sticky burs or other devices for catching in the coats of animals were found.
It is among the Compositæ or daisy family that the wind is seen to work most effectively as distributing agents of its light-plumed fruits. In the Falkland Islands, St. Helena, and Prince’s Island, all from 300 to 1,500 miles from the nearest land, species of Senecio or groundsel have been found that may well have been wind-driven onto these remote islands from the mainland. In fact the whole genus Senecio, consisting of over two thousand species, and of world-wide distribution, has in all probability been spread largely by its wind-borne achenes. They are not edible, nor are they suited to carriage by the ocean currents, but some of them are known to stick in the plumage of birds.
But it is hardly necessary to cite these far-flung examples of the wind’s action in distributing seeds, for there are many interesting cases much closer home. The tumbleweeds, such as the false indigo and certain grasses, are familiar sights scudding before the wind over prairies in the West and open places in the East. By a kind of foresight the winged seeds of the pines are so weighted, due to lack of symmetry, that instead of sailing quickly to the ground, they tumble and flutter about, thus prolonging the time of flight. And in the linden the curious winglike attachment from which the stalk of the fruit arises is admirably fitted to slide over the snow and ice upon which, through their often tardy falling, they are deposited. Scores of such adaptations of structure to function are known, where there seems direct response to conditions and there is the temptation to say that such adaptations are caused by the wind or other agency. Nothing could be farther from the truth, as plants do not produce winged seeds or luscious fruits to insure seed dispersal, but the seeds of plants having such devices, from whatever cause, are naturally favored in the ceaseless struggle to occupy new land, which is quite another thing.
WATER
The scattering of seeds along streams is too common a process to need more than a mere mention here, for it is to be seen along any fresh water stream at harvest time. But seeds that may be carried by ocean currents have a much greater influence on the distribution of plants, such as the coconut palm, already mentioned among the food plants, and now common throughout the tropical world.
The number of seeds that will float in sea water and still keep the power of germinating is not very great. Most seeds sink at once; many will float for months, but are useless when they reach land; but those that will both float and grow afterward have worked some curious changes in the floras of different islands. Sometimes the great ocean currents, like the Gulf Stream, appear particularly futile in the fruits they carry such long distances, as the pods of a tropical vine from the West Indies are not infrequently found on the coast of Norway and even of Nova Zembla, of course uselessly. But many ocean currents, particularly in the Pacific, have carried fruits and seeds thousands of miles, they have even carried so-called floating islands of vegetation bodily.
Perhaps the most remarkable case is that of Entada scandens, a tropical vine of the pea family, bearing large pods, sometimes several feet long. The plant is not typically a seaside plant, and there is evidence that fruits matured in the shade of its usual forest home will not float. Those that grow nearer the coast and more in the open develop, through partial drying, a small air chamber inside upon which the seed depends for buoyancy. Its original home is apparently somewhere in Central America, from the west coast of which it spread over the Pacific to the shores of the Indian Ocean. Over the Atlantic it has reached the shores of tropical Africa, and in fact wherever ocean currents cast up their refuse on lonely beaches parts of the giant pod or individual seeds of Entada scandens are found. Partly fossilized remains of them have been taken from peat bogs along the coast of Norway, of course dating since glacial times, but showing by their presence there how long this water-borne seed of the pea family has been attempting to populate the earth. Of course all that do not reach tropic shores are lost, but nearly throughout the Pacific Islands, with some exceptions, the vine is now established. The extraordinary feature of it all is that scarcely half the seeds of the plant will float at all, nearly all inland forms sinking at once in the fresh water rivers into which they may chance to fall. Only those that grow near the sea, in mangrove swamps and the like, or at any rate near brackish water, will float. These, however, apparently float indefinitely without loss of germinating power. There is abundant evidence that many plants of oceanic islands have similar characteristics so far as their inland and seaside forms are concerned.
While Entada scandens has spread in spite of unfavorable adaptation for seed floating, there are some plants whose seeds always float, and in spite of the sea water retain their power of germination. H. B. Guppy, from whom much of the above data are taken and who has experimented for years on the buoyancy and germinating power of sea-borne seeds, reckons about two hundred species that may have spread by ocean currents. Many of these are nearly world-wide in their distribution within their climatic requirements, and most are confined to the tropics.
2. Invasion, Migration, and Relics
It is perhaps a natural enough question why such elaborate and effective methods of seed dispersal are necessary and why plants, once they grow in any particular locality are not satisfied to stay there. The answer to this is that individual and racial competition is so great that without means of dispersal, which may be looked on as equipment for seeking a more favorable site, species would often be crowded out.
There is no better place to see this than at the edge of a forest and grassland. The presence of the forest tells us at once that what may be described as the forest type of climate must have existed in the past to have produced the woods. In clearing off parts of this forest the openings will usually be grassland at first, but never in the end if nature is allowed to work out the solution. Along the edge of such a forest will be found a host of pioneers pushing out among the grasses, making ready, by conditions of shade, protection from drying winds and other influences, for the seedlings of the forest trees that creep slowly but resistlessly out to capture areas that by right of previous occupancy belong to them. In practically all parts of the world, with a few local exceptions, wherever the forest was the original type there is this ceaseless struggle to reclaim the open places, often or usually peopled by grass. It may be set down as almost a rule that if the open places produce herbs with broader leaves than grasses, the forest will capture the area many years sooner than if grass alone is the temporary tenant. Grass by its exclusive growth, its complete monopolization, so far as low seedlings are concerned, of much light and nearly all available surface water, is singularly well able to take care of itself once it is thoroughly established. But plotted and marked areas of this contact between forest and artificial clearings in it, show that in the end the forest will win, often at the rate of five hundred feet in a hundred years, sometimes much quicker than this.
If, on the other hand, what may be called the prairie type of climate has resulted in the formation of grassland, which has happened in our own West, in the steppes of Russia and less extensively in many other places in the world, forests can hardly ever get a foothold. Where, as in river bottoms, they sometimes flourish, the line between forest and grassland is sharp and apparently an impassable barrier for trees.
This invasion of immediately adjoining territory is going on constantly not only by different types of vegetation but by the units of it. The frequency of different plants in different years, their final ascendancy or extinction, all point to the struggle for expansion which in a score of ways the plant world is constantly waging. In many cases we are not yet able to see the struggle, but only its results, while in some places the bitterness of it may be gauged by the dead and dying that strew these silent battle fields.
The dominance of certain species of plants, such as grasses on a prairie, the fir and spruce in the coniferous forests of our North, the blue-gum trees in parts of Australia and the giant dipterocarp forests in the Philippines, are all based on the ability of individuals to spread from their point of origin. All species of plants must have one day been of very local distribution and confined to the region where they were born, but from their often very modest beginnings some of them at any rate have gone to the ends of the earth. The common bracken fern is found in nearly every country in the world from far northward through the tropics to the antipodes, and yet no one knows where its original home might have been. Somewhere up in the northern Andes it is supposed that the first ancestor of the huge daisy family had its origin. From there its many descendants, now ramified into hundreds of genera and over eleven thousand species, have spread to the very limits of plant growth. And the daisy family is one of the most recent of all the families of plants.
The distribution of plants is of many kinds depending on local conditions of climate and soil, on individual and racial competition, on methods of fertilization or other means of propagation or seed dispersal, and particularly upon the distribution of the plant or its ancestors in past ages. While it is often difficult or impossible to determine upon which of these factors, or upon what combination of them the distribution of any particular species is based, certain facts of plant dispersal appear to be indicated by a study of existing floras. A few instances must suffice here to illustrate the principles by which many plants are scattered over the earth, or else restricted to localized regions, and which, without knowledge of the factors involved, seem merely the wayward caprice of nature.
In the flora of eastern North America there are many genera that appear to be endemic there (found nowhere else), but are actually duplicated in eastern Asia, if not as to species, at any rate by plants so closely allied as to be of obviously common origin. These plants are unknown in Europe, or on our own western coast. The skunk cabbage, sassafras, twinleaf, May apple, Canada moonseed, spice-bush, ginseng, sour gum, trailing arbutus, fringe tree, lopseed and many others, all fairly common in eastern North America, are unknown between this and eastern Asia where, if not the identical species, which often happens, closely related forms are duplicated. The explanation of such discontinuous present distribution appears to be that at some time in the distant past there was a land connection between Asia and the western coast of America, the remnants of which form the Aleutian Islands, and over which there was a constant migration of plants and animals. With subsequently changing climatic conditions on our own western coast, due to warm ocean currents, most of those Asiatic migrants, or it may have been a migration in the opposite direction, were crowded out by later types which now dominate the Pacific coast. There is small chance that these plants were spread by birds, as an east-west bird migration is hardly likely. Nor is there any record that the seed of these plants, even where they might float, would survive ocean transport.
Sometimes the ancestors of now widely separated species or genera once covered all or nearly all the intervening area, and again sometimes only a minute fraction of the ancient distribution is left at the present time. Our own Big Trees of California were once known to grow in England, Iceland, all through central Europe and eastern Asia, Australia, New Zealand, southern Chile, and from Texas to Alaska. In the face of such widespread occurrence in the past their present distribution over an area of a few square miles is merely a pitiful relic of ancient grandeur. Scores of cases are known where, instead of a single outpost as in the Big Trees, there are only a few widely scattered survivors from a probably much more continuous distribution in the past. In northeastern North America there grows along pond sides and fresh-water beaches the shore-weed, a relative of our common weedy plantain. The only other species, and a close relative, comes from the southern tip of South America.
Many other species now existing are to be viewed only as relics of a bygone, often much more continuous and more widespread dispersal. In the persimmons, of which over a hundred different kinds are now known, the original distribution covered all North America, all Europe and Asia, except the Scandinavian Peninsula, all of Africa, northern Australia and South America. To-day the genus is restricted mostly to tropical Asia, southern Africa,

Some of the Commoner Plants of the Desert in the Southwest. The fanlike branches at the left are the ocotillo (Fouquieria), the two short tree cacti and choya cactus (Opuntia) and the leafless tree in the central background the palo verde (Parkinsonia). (Photo by the late Edward L. Morris, released for publication here by the Brooklyn Museum.)

Grassland and Tree Vegetation. All over the world there is a contest between grassland and tree vegetation for dominance wherever both occur. In this thorn veld in Natal the struggle is particularly keen. (After Bews. Courtesy of Brooklyn Botanic Garden.)
northern South America, and in North America it has dwindled to a handful of species confined to the region south of the Great Lakes and generally east of the Mississippi. In Mexico and south of it the genus is better represented. But with the persimmon, as in so many other types, our existing species are the remnants of preexisting ones, without a knowledge of which their present dispersal would be impossible to explain.
It is easy to reason from the foregoing that widely separated but related species of plants are all either very ancient, or directly descended from ancient ancestors, and that other things being equal a widely dispersed species is older than one with a restricted distribution. It is most certainly true that relics are unquestionably very ancient, and do actually represent the last outposts of a preexisting condition. But many isolated plants are relatively very new, so far as their immediate origin is concerned, as witness the hosts of young species of the daisy family, some of which have spread scarcely at all from their obvious point of origin. In judging of the distribution of even the commonest tree or shrub of our woodlands there are these links with the past, as well as response to present conditions, to be weighed if we are to understand the story aright.
Once a plant reaches a new and for it a strange country it is remarkable how quickly it will often capture the new territory. In the United States over six hundred of our commonest weeds have come from Europe and Asia. The daisy, dandelion, wild carrot, many hawkweeds, dozens of wild mustards, and many others are among the somewhat undesirable immigrants that now reach over a great part of the country—all brought over by the early settlers of America. From their home in subtropical Asia the lemon, lime, and orange have invaded every part of the tropical world. Once, in the most remote part of the Sierra Maestra Mountains in eastern Cuba, where only by the most arduous cutting could a passage through the dense tropical forest be forced, the writer found within a few square rods an orange and a tree from tropical India. Such cases could be multiplied, and all over the world we see this endless struggle of plants to conquer new territory, often at the expense of existing vegetation. On Long Island, New York, the introduced locust tree, brought from the southeastern United States about a hundred years ago, has completely routed the native trees in many places along the north shore of the island. And in Hawaii, some seeds of a screw pine, washed up among refuse along the beach or brought in by early aborigines, have made this Malayan plant a common tree thousands of miles from its home.
3. Home Economy of Plants, or Ecology
The geographic distribution of species of plants may be, as we have seen, the result of the geological changes of the past, of bird migrations, of more or less fortuitously water-borne seeds, or more usually of the slow spreading by invasion of those species apparently not so well supplied with external helps to dispersal. But no matter where plants grow, nor how they got there, they must fit the particular environment in which they find themselves or perish. This home economy of plants, or how they meet the environment and each other, is called Plant Ecology, a phase of botany now much studied, for it tells us more directly than most other plant research what the actual response to various factors of the environment may be. Just as plant distribution is the reflection of many, usually widely operating forces, so ecology narrows down to individual plants or groups of them the impact of the immediately surrounding conditions upon vegetation.
The basis for all study of the response of plants to the conditions under which they grow must rest upon the response of their different organs to those factors, just as our general movements are dictated by sufficient food or air or water to keep ordinary bodily functions going in the ordinary way. But the study of such plant response has shown that certain kinds of environmental conditions have resulted in quite similar response nearly throughout the world. Often totally unrelated plants assume characteristically similar growth forms where the conditions in widely separated areas are climatically or otherwise similar. In our own Southwest we have the dominant cactus vegetation, matched in parts of South Africa by giant cactuslike spurges. In Mexico we find the wealth of century plants, which are confined to the New World, matched in the Old World dry regions by the aloe, a group of succulent plants nearly as well suited to such areas. The species of plants characterizing peculiar regions may well be the result of geographic distribution that rests on more widely operating factors such as we saw in the previous section of this chapter, but the type of plants growing in a particular place hardly ever fails to be dictated by the local condition. With this in mind, a vast amount of time has been spent in studying the various factors of the environment, such as climate, soils, altitude, light, etc. And an equally valuable study has been the response of individual plants or their organs to such conditions. From this great body of information, obviously impossible to include here, we all recognize certain well-marked societies or groupings of plants which, wherever they occur, exhibit similarity of general response to the different conditions responsible for their occurrence. Once these typical plant societies or groupings are understood we can recognize them wherever they may occur, and we shall see that they are as widespread as are plants themselves.
Just as societies or races of men have often obscure beginnings, reach a climax, and afterward die, so these plant societies may be considered as exhibiting a similar progression. What these plant societies are, at least the more important of them, will be considered in the next section of this chapter. It should never be forgotten that the species of plants making up the dominant plant societies in different parts of the world are dictated by quite other conditions than those that result in the dominance of the society itself. Perhaps as good an illustration as any is the aristocratic type of mankind, recognizable throughout the world by the possession of finer qualities than the common run, but differing in individuals as much as the best type of Americans, the British peerage, and the samurai of Japan differ one from the other.
Not the least interesting feature of these plant societies is that we must view them as associations of plants, often of widely differing origin due to the vicissitudes of plant distribution, but all taking their part in the society to which they belong and often, as individuals, losing their life that the society may live. Upon such a conception a wood or prairie, or river bank, or salt marsh or alpine garden upon a mountain summit are, with many other plant societies, places of intense conflict. More cruel than any human society, these plant communities exist under conditions where only the individually strong survive, and only those societies are destined to reach their climax which can take advantage of every aid, quite without regard for severe losses or even death to the individual members of it. It is as if we poured into a crucible molten metals from many different sources, and after the incredible and relentless forces of manufacture had worked their magic upon them there resulted a product, purified and cleared of all dross. So the inexorable and relentless processes of nature work over the materials found in these plant communities, the results of which are the dominant types of vegetation in the world to-day. With this understanding of the part they play in plant distribution we may now consider a few of the most widely recognized plant societies and see how they have affected the vegetation, sometimes even the history of the regions in which they are found.
4. Some of the Chief Plant Societies
Forests
No one who has ever seen both our temperate forests and those in the tropics can fail to be impressed with the difference between them. Not only for the different plants in them, but for their wholly different aspect, tropical and temperate forests stand far apart as an expression of the forest covering the earth. Not all of us realize, however, that the heat of the tropics is not the deciding factor in the luxuriance of those dim jungles, and that a rainfall far above anything occurring in the United States is even more important. Upon the distribution of rainfall depends the occurrence not only of the two forest types that will be mentioned here, but of most of the other chief plant societies.
TROPICAL RAIN FOREST
A small section along the lower side of the Gulf of Mexico, the northeastern edges of Cuba and Santo Domingo, nearly all of the region drained by the Orinoco and Amazon Rivers; in the Congo, Zanzibar, and Madagascar in Africa; all of southeastern Asia, including the East Indies and part of the north coast of Australia—these comprise the regions of the tropical rain forest. All of them have, besides continuous heat, a nearly continuous or in some places a periodic rainfall, averaging over, and usually much over, eighty-five inches a year, as compared with about half that near New York. There are, of course, other places in the world where these rain forests, so called from their abundant moisture and some of the effects of it, are found. But in the regions mentioned they are at once the most wonderful and to most white men the most awesome manifestations of the plant world.
Such forests seem, and actually are, pulsating with life, as instruments stationed in them have many times proved. With some kinds of bamboo growing over two feet a day, and a eucalyptus tree in Java forty-five feet in three years—and these are not isolated cases—the tremendous annual increase in the amount of vegetation can be glimpsed. Of course not all the plants in them grow at any such rate, but the great heat and abundant moisture does make tropical rain forests irresistible in their power. Plantation owners, and railways that have been run through such forests, wage constant warfare against the recapture by the teeming forest of man’s intrusion of it. The writer once saw in Santo Domingo a railway cut through such a jungle and abandoned only two or three years before. Not a trace of the roadbed could be found, ties, rails, and switches all covered with a dense vegetation, and overhead the canopy of the forest had closed over the opening and was already sending down hundreds of adventitious roots that would complete the obliteration of man’s handiwork. Everywhere there is the evidence of vegetable life run riot, ever crowding and pushing to close up openings made by the crashing down of old trees or the artificial clearings of man. Those living on the edges of such forests speak and think of them as dim, mysterious places where strange creatures and the ever-present fevers join forces with the vegetation to keep out humankind. That they are places of actual danger everyone knows who recalls that Stanley’s trip through equatorial Africa cost one hundred and seventy lives, many of which were sacrificed to disease and strangely enough nine were lost through starvation. While the tropics supply much of the food used there, these jungles produce almost none of it and because of the scarcity of edible fruits, the extraordinary difficulty of getting about and collecting what does grow, starvation faces anyone who goes into them without adequate supplies.
In the Amazon grows the largest water lily in the world, Victoria regia, with giant leaves upon which a moderate-sized man may stand in safety. It produces a flower over a foot in diameter, and it is surrounded by a forest the like of which it is difficult to describe. H. H. Rusby, who spent two years in this region hunting for medicinal plants, has described the country a few hundred miles below where Victoria was discovered. He writes: “Passing down the river Madeira to the lower Amazon, we come into a region of such grandeur in its vegetation that it is difficult of comprehension, even by one who is an eyewitness. Everything is in such proportion that one is apt in its size to miss the gigantic. Many of the trees of this region are undoubtedly many centuries old and appear to be good for many centuries more. Most of them have enormous buttresses at the base, and these buttresses often begin as high above the ground as are the tops of ordinary forest trees in our land. All are bound together with an impenetrable mass of tough vines. Running through these swamps are the most beautiful little bayous or canals. Nothing can exceed in interest and delight a day’s canoeing among these narrow waterways, although there is great danger that the inexperienced boatman will hopelessly lose his way. In the rainy season this river rises sixty feet or more above its low water mark and the boatman travels among the tree tops which a few weeks before were high above his head.”
The abundant water supply in the rain forests results in an atmosphere saturated with water vapor and in some of them it is a common sight in the morning to see the forest rising out of an unbroken blanket of mist. As this dries up under the heat of the day, or if there occurs one of the torrential downpours to which such regions are frequently subjected, there rises from the forest in plainly visible waves a vast quantity of water vapor. It is this that has so often made them be described as steaming forests. The water requirements of the plants are more than supplied, nay, there is such a surfeit of available water in all these forests, that there are numberless devices to get rid of the excess. Dripping points to the leaves, already described in an earlier chapter, are common. But in addition many plants have wonderfully colored leaves such as Begonias, some relatives of our jack-in-the-pulpit belonging to the Arum family, many orchids, and other plants. The colored leaves in the predominantly dark green and gloomy rain forest, because of their greater absorption of light and consequently higher transpiration rate, are of decided advantage.
While there is thus very little or in fact almost no struggle for water in the rain forest, the struggle for light is intense. In the deepest and most luxuriant of them the gloom of the forest floor is notorious and it was by no means a figure of speech for Stanley to describe his trip through equatorial Africa as “Through the Dark Continent.” So dark are most rain forests, and, as we have seen in a previous chapter, so inexorable are the plants’ demands for light, that the various devices to insure it are perhaps the one great difference between these forests and those of temperate regions. One effect of the struggle for light is the enormous production of vines often running hundreds of feet through the tree tops. In India the Calamus, or rattan palms, with stems no thicker than a walking-stick, will completely interlace the foliage of the canopy. Thousands of slender whiplike roots and stems of such plants descend from the topmost heights of the forest canopy, where the plants to which they are attached make such an inextricable tangle among the tree tops that orchid collectors have been known to travel considerable distances over the matted vegetation, with, it must be confessed, considerable danger. These vines or lianes as they are called, are however, often as thick as a man’s body and armed with great hooked prickles, an obvious aid in catching some support to reach that essential light for which all plants in such places are ever striving.
Besides the bewildering tangle caused by these lianes, the rain forest is further impeded by hosts of epiphytes or plants that are mechanically attached to tree trunks, branches, or anything else that will raise them to the light. Of all the plants of such regions the epiphytes are the most light-demanding. They must not be mistaken for parasites, as they have roots of their own through which they absorb nourishment, mostly as water vapor, but also as liquid water held in the bark and refuse in which they grow. Thousands of orchids are epiphytes, also ferns, and, only in the American tropics, thousands of different relatives of the pineapple. Many of the latter are among the most gorgeously colored of all plants, their superb foliage being much sought after and the specimens largely grown in our greenhouses. In most rain forests every available inch of space is covered by these epiphytes, so that no bark, scarcely any branches, are to be seen but those clothed in this motley array of plants that use the support to get the utmost possible light. Many of these epiphytes have rosettes of leaves arranged for holding water, and after a sharp thunder shower followed by fresh wind the writer has seen the ground strewn with thousands of relatives of the pineapple which, with the added supply of water, were unable to stand the strain and were consequently wrenched from their lofty perch. So enormous is the combined weight of these epiphytes, together with the lianes, that many trees crash down under the strain long before their time. Perhaps no sight of the rain forest so convinces one of the struggle for light as to see one of these forest monarchs come crashing down loaded with thousands of plants that have been using it for support, and to escape which it has pushed its canopy to the utmost limits of its growth. Such contests are common in a forest of which only the barest outlines can be conveyed to those who have never seen it. To those who have had that good fortune any description palls beside the wonderful actuality.
It is scarcely to be wondered at that these steaming rain forests with their gloom, and, as they were once described, “all hung about with fever trees,” should be dreaded by many, and the subject of fabulous tales to the credulous. The almost incredible difficulty of getting through them, not to mention the savage animals that inhabit many of them, have not lessened the tendency to exaggerate about these great forests. But the truth about them is so far beyond belief, the strange plants that intrepid explorers have brought out of them so almost incredible, that it only excited a temporary wonderment when the largest flower in the world was discovered in such a forest in the Malayan Archipelago.
Sir Stanford Raffles and Dr. Arnold, while exploring in Sumatra during the year 1818, discovered what was called “the greatest prodigy of the vegetable world,” and no flower since found equals its size. The plant, without stem or leaves, consists wholly of one gigantic flower about nine feet in circumference, and was subsequently called Rafflesia Arnoldii. It aroused a sensation in England which was not abated by knowledge of the fact that the flower is a parasite on the stems of certain tropical plants related to the grape. That such a huge flower should be the product of a parasitic mode of life is one more illustration of how this and related irregularities occur in widely separated families of plants, and under varying conditions. Relatives of it have since been found in India, some parasitic on roots, others, as in Rafflesia Arnoldii, on the stems of vines. The sticky seeds are in all probability carried from place to place on the hoofs of elephants, to which they have been known to cling. Only if they are deposited on a bruised or otherwise exposed tissue of their future host can they grow. These curious plants have been actually cultivated in the greatest tropical botanical garden in the world, at Buitenzorg, Java.
The original collectors of Rafflesia Arnoldii could scarcely credit their senses when they saw for the first time this extraordinary plant, whose whole life is spent in producing this great flower and fruit. As one of them says: “Had I been alone, and had there been no witnesses, I think I should have been fearful of mentioning the dimensions of this flower, so much does it exceed every flower I have ever seen or heard of.” The odor of the flower is repulsive, and, with its great size and curious mode of growth in the dark rain forest, it is surely one of the strangest productions of the vegetable world.
But as the utmost development of the plant world, and producing the greatest profusion and richness of plant life, these rain forests are, beyond the sporadic occurrence of such wonders as Rafflesia and some others, places of extraordinary interest. With every inch of space occupied by plants, the very epiphytes often having on their leaves still smaller plants, we see here what nature will produce when the maximum conditions for plant growth are so nearly perfect. Theodore Roosevelt in his book, “Through the Brazilian Wilderness,” gives a vivid picture of the rain forest there, and it may well end our account of those most interesting of all plant societies:
“In one grove the fig trees were killing the palms, just as in Africa they kill the sandalwood trees. In the gloom of this grove there were no flowers, no bushes; the air was heavy; the ground was brown with moldering leaves. Almost every palm was serving as a prop for a fig tree. The fig trees were in every stage of growth. The youngest ones merely ran up the palms as vines. In the next state the vine had thickened and was sending out shoots, wrapping the palm stem in a deadly hold.
“Some of the shoots were thrown round the stem like the tentacles of an immense cuttlefish. Others looked like claws that were hooked into every crevice, and round every projection. In the stage beyond this the palm had been killed, and its dead carcass appeared between the big, winding vine trunks; and later the palm had disappeared and the vines had united in a great fig tree. Water stood in black pools at the foot of the murdered trees, and of the trees that had murdered them. There was something sinister and evil in the dark stillness of the grove; it seemed as if sentient beings had writhed themselves round and were strangling other sentient beings.”
Many other forests in the tropics, where the rainfall is less, or less regularly distributed, are not unlike our own, having rather regular periods of leaf-fall that come with the dry season rather than with the autumn. The trees are of course never the same as ours, but the general aspect is not very different from that of temperate forests.
TEMPERATE FORESTS IN AMERICA
The transition from the tropical rain forest to our own woodlands is one of the most interesting, as it is certainly the most gradual in nature. Lack of space prevents our stopping to note those strategic points along this pathway from a hot, steaming forest to the cool shade of our open woods, where traveling, in at least a virgin forest, may be done easily on horseback. As we come northward, and if we could travel continuously through the forest, we should lose first the epiphytes, then most of the lianes, and finally all the condition of vegetation crowding into every inch of space suitable for it. While trees in our virgin forests are as thick as they can be, the forest floor is open and on it grow only a few herbs that will stand lack of sunshine.
But the really great difference is the long, unfavorable season in temperate regions where the forest must drop all its leaves, after, in at least our own Eastern States, the most gorgeous foliage coloring of any forest in the world. The winter months when the woody vegetation is practically fully exposed to the elements, are particularly severe in their effects. Leaf-fall, which is such a common sight as to arouse scarcely any interest, is the only device by which the great bulk of our forest trees survive, and only in the southern part of the region are there found such woody plants as the mountain laurel, rhododendron, American holly, and a few others which are evergreen but not cone-bearing ones, and are the only reminders of the truly evergreen forests of the tropics. The winter winds farther north and in the central treeless part of the United States prove too much for many kinds of trees, for instance, all the oaks and sassafras, none of which go very far north.
There is sufficient rainfall to produce forests much farther north than they are found, but lower temperatures prevent trees from growing just as too little water stops their growth altogether. Toward the northern limits, or upon high mountains, the upper limits of the forest, we get the best idea of how persistent woody vegetation is in the general forest area of the eastern United States. Stunted, wind-swept and weather-beaten trees are often found only a couple of feet high and over sixty years old. Sometimes they will be flattened out on the ground or on bare rock, making great patches of bushy growth quite unlike their lofty relations in the lowlands. The growth rate for such plants is so slow that their annual rings are all but obscured. With such persistence in the production of these elfin forests, high up on mountains under the most unfavorable conditions, it is little wonder that below this are trackless woods, and that the northeastern United States has one of the finest developments of the temperate or summer forest in the world.
Nor are all our woods of this general type made up of the same species, for everyone knows about the endless spruce and fir forests of the north, exclusively evergreen, and in the summer nearly always moist. This spruce belt stretches practically across the continent, where, in the West, other and our most gigantic evergreens, replace the eastern spruce and fir. A little farther south is the region of the white pine now nearly unknown as a virgin forest type, as its great value led to early and ruthless cutting. The white pine region is generally the area from New England southward along the Alleghenies and westward to Illinois. But the most characteristic of the temperate forest types is our summer forest, so called from its general lack of evergreens and its beautiful green foliage of summer and its bare branches in winter. Beech and birch and maple, in different proportions according to local conditions, predominate in such woods. These hard-wooded trees, with many others that are scattered through them, have been among the most valuable of all the natural plant products of our country and their destruction has been upon such a scale that only in a few places may the virgin forest be seen at the present time. Where it does occur we find the forest floor often with nothing growing on it except a mass of spring flowers which are half matured before the leaves of the forest canopy close out nearly all the light and much water and put them to rest until another year. The great preponderance of spring-flowering herbs in Eastern North America is due to their early warming up before the foliage of the trees cuts off their light. And in some virgin forests of this sort, particularly where there is a large mixture of oak, the writer has seen hundreds of square rods without undergrowth or herbaceous vegetation of any kind. Such places, very rare indeed at the present time due to senseless and wicked cutting, are rather dark, perfectly open to view for hundreds of feet ahead, and dotted only with the huge trunks of the trees that characterize this climax type of the temperate or summer forest.
The absence of direct sunlight and interception of much rain under the forest canopy has other effects besides stopping the growth of herbs and shrubs which are common enough along the edges, or where openings are made by the fall of old trees. It prevents the germination and growth of nearly all the seeds falling from such trees, and in a really virgin forest of this sort, almost no seedlings will be found. Upon cleared or open land thousands of saplings will cover much of the ground, but nearly all these will die off due to crowding, and leave as the climax only enough trees to close over the forest canopy.
Forests may be found in all stages of succession from those just beginning the process to those final forest monarchs which, having won out in the race, are, until one of them falls, often slow to perpetuate the type. For, as often as not, a new growth will spring up once a very large tree falls, and a very different kind of growth from the climax forest. At once a host of species, that one might almost say had been waiting for the tragedy of the monarch’s fall, will rush in and convert the opening into a nondescript brush patch, out of which will rise another tree that means business. As it grows to maturity, it kills off these smaller triflers one by one, until, when the canopy is finally closed, all of them will have disappeared, or, as often happens, retreated to other parts of the forest, where they wait for another chance. This succession of different kinds of growth in a temperate forest is so well known that in England they have for centuries practiced it, for commercial or pleasure purposes. In their oak-hazel copses they cut the trees enough to partly open the canopy, which permits a dense growth of hazel bushes and other plants. Every twelve or fifteen years the latter are cut down for various purposes, and will gradually spring up again to renew the dense growth. The spacing of the trees is sufficient for them to branch freely and yet not close the canopy enough to kill off the hazel. The trees are cut off a few at a time, not oftener than one or two hundred years in any one spot. By this procedure the owner gets a regular crop of hazel once in twelve or fifteen years, occasional big trees, and on many places a cover for pheasants. Under the hazel there is a regular progression of herbs, very plentiful just after the bushes are cut, and decreasing almost to nothing when the end of the growth period of the bushes is near.
The English oak-hazel copse, now much less grown than formerly, and the general lack of undergrowth in our own virgin forests, are both responses of the forest to light and other factors that are related to an open or closed canopy. As we stated a page or two back, it is not impossible, it is even frequent in some parts of the country, for a forest to produce, by its own growth, conditions inimical to its perpetuation. Where the casual falling of a forest giant is the only opportunity which that forest offers to perpetuate its type, it may well be said to be a climax forest, incapable of further development. But in some such woods a curious provision of nature insures an invasion of the gloomy forests by trees less light-demanding than the dominant ones. And often these trees that can get along with less light will capture considerable parts of forests that light-demanding trees could never conquer.
If the soil in which these temperate forests happen to grow is sandy or otherwise poor in plant food, the broad-leaved trees that make our woods such a delight in summer are replaced almost universally by pines. Along the sandy stretches of the coastal plain from Long Island, New York, to the Gulf, there are immense tracts of these pine forests, different species often being locally dominant, such as the pitch pine in the pine barrens of New Jersey and the long-leaf pine farther south. Almost throughout the world there is this monopolization of the poorer and drier forest sites by pines, which maintain the forest plant society in regions where the broad-leaved rapidly transpiring trees could not grow.
No account of forests, however brief, can omit some mention of the greatest agency for their protection in North America, the United States Forest Service. With corporation and individual cutting and attendant fire hazard upon a scale almost beyond belief in its ruthless disregard of our chief natural plant product, the Government soon found that Federal ownership or control of forests was the only policy that would maintain even a partially adequate timber supply. National forests, set aside either for pleasure or profit, now total more than the area of France or than all the New England and most of the Middle Atlantic States. These huge tracts, in every part of the country where forests are found, are well managed, properly planted, and most important of all, constantly guarded against fire. Forest fires not only destroyed over $25,000,000 worth of timber annually, but leaving devastation behind them, depleted the water supply in many parts of the country. Nothing but forests will hold the rainfall, to release it slowly through a thousand rivulets and springs that are the source of countless rivers. With the forest cut or burned off these streams are dry most of the summer and raging torrents for a few weeks in the spring, washing out all the priceless accumulation of the ages which the forest has conserved for its own and our benefit. While the reservation of these great national forests has worked individual hardship, experience for many years back in India and Germany shows Federal ownership or control the only wise policy.
Forest covering, whether temperate or tropical, depends for its occurrence all over the world upon an adequate rainfall. As we have seen in the tropics, this may be so great that coupled with the heat it produces a wealth of vegetation beyond the powers of description. Where it is less and the country cooler, the forests are of a different type, but even there the forest covering is, without interference, practically complete. Where, as in parts of Chile, southeastern Australia, and of Japan, there is a heavy rainfall but cool climate, there is a so-called temperate rain forest. Such forests are cold, drab, wet woods of peculiar aspect and extreme interest. For in them grow trees sometimes related to our own, but, due to the special conditions, producing a forest landscape quite unlike anything in America. It would seem as if we might almost plot the distribution of forests in our own country with a weather map showing rainfall, and such is actually the case. When the rainfall becomes less than will maintain forest growth it stops, often very abruptly. Generally speaking, the region west of the Mississippi, and some just east of it, westward to the mountains, is entirely devoid of forest, except in the river valleys. The forests give place to an entirely different type of vegetation—the prairie or grassland.
THE PRAIRIE OR GRASSLANDS
In nearly every region in the world there is an absence of forests and a replacement of them by grasslands, where the rainfall is less than about twenty-five inches a year, and where the winter winds, often far below the freezing point, are hostile to trees. The distribution of the rain mostly through the growing season also makes a condition peculiarly unfavorable for trees during the winter. Someone has said that the nations have fought since the days of the Romans for the belt of grassland which these climatic conditions have produced all round the world, and it is certainly true that these naturally grass-covered areas have produced the cereals of the world, all of which, except rice, grow to best advantage in such regions.
With of course different species of grass and quite different associated herbs, these grasslands are now found in our own prairies, the steppes of Russia, the plains of Hungary, the pampas of southern South America, the grasslands of Australia, the veld in Natal and in many other but mostly less extensive developments. Some of the grassland regions are warm, but without more rainfall than characterizes such areas the greater heat does not produce a forest. Usually, but not always, these grasslands are not found near the coast, where, as in America, the rainfall is double that of the plains and produces the forests that clothe the Atlantic and Pacific sections of the country. From the east westward there is a gradual decrease in the rainfall, until from about the Mississippi to the mountains it falls below the point where trees can compete with the prairie.
Another characteristic of prairies that once they have started tends to keep out trees is their almost annual firing. Tree seedlings cannot survive this, and we know that the Indians fired huge tracts of prairie every year, not to mention fires started by lightning which may set fire to grasslands and actually does set fire to forests every year.
The prairies in the United States—perhaps the most extensive in the world—are characterized chiefly by several grasses, buffalo grass (Buchloë dactyloides), gama grass (Bouletoua oligostachya), and several prairie grasses, such as Sporobolus asperifolius, Koeleria cristata, and some others. Among these, depending on the soil, are hundreds of prairie flowers which, during different parts of a single season, give quite different aspects to the region. Both the grasses and their associated herbs are well protected against too violent transpiration which their exposure to nearly continuous sunshine, high summer heat, and very considerable winds makes particularly active. In many places where the country is rolling, the lower and moister sites, besides developing more luxuriant growth of prairie plants, permits low shrubs, and in river bottoms even trees to flourish. But climatic conditions of small rainfall, high winds, and bitter winters make anything like a forest development out of the question.
In some regions, both in America and central Europe, a rainfall that is high enough to permit trees to exist and low enough to favor at the same time a grassland formation has resulted in the parklike landscape that creates the most beautiful scenic features of the regions where it occurs. In such places there are irregular patches of forest and grassland, and the struggle for supremacy as between the different types depends, not upon the general climatic conditions to which they both respond, but to local conditions of available water supply and soil conditions and often upon fires. Naturally such regions are places of intense strife for dominance, and in them some remarkable collections of plants have been found.
One of the most interesting of these struggles between grassland and woody vegetation in a region climatically able to produce both, is in Natal. Large sections of that country are grasslands or veld, as the people there call it. Scattered through the veld are various species of acacia trees, locally called thorn, with feathery compound leaves. These do not shut out enough light to prevent the development of grasses directly under their shade, yet the annual firing of the veld prevents, except accidentally, the production of the acacias. But the seeds of this tree, whether from long usage to this burning or not, are actually hastened in their germination by the firing, and it is a common practice in that country to roast or partly boil the seeds of the tree to hasten germination. The presence of the climatically favorable environment for both trees and grassland results in the latter being the dominant type of vegetation over large tracts of the country, largely because fires destroy tree seedlings, and yet the tree seeds, by a quite extraordinary fitness for their peculiar environment, offer a measure of insurance against the total destruction of woody vegetation by the grassland.
The pampas of the Argentine have been vividly described by P. G. Lorentz, who, in writing of the drier parts of it, says: “Viewed from a distance, these grasses seem to form a close grassy covering, and the pampa presents the appearance of extensive grassy tracts whose coloring varies with the seasons: coal black in the spring, when the old grass has been burned; bright green, the color of the mature grass; finally—at flowering time—when the silvery white spikes overtop the grass, over wide tracts it seems like a rolling, waving sea of liquid silver.
“After the Gramineæ (grasses), the family of plants that is represented in the pampas by the greatest number of individuals is that of Compositæ (daisy family); usually twiggy undershrubs with inconspicuous flowers, a bright yellow Solidago (golden-rod) alone gleams out from among the others.”
Here, as in the other grasslands of the world, if a local water supply above the general requirements of the grass exists, there is always a small element of woody plants, low, thick-leaved shrubs usually, but where water is more plentiful, trees, as in the region in Natal, already mentioned, and in many other parts of the world, notably parts of Australia, China, Brazil, and many sections of the western part of the United States.
We see in grasslands a plant response to rainfall and other climatic conditions which, with a little more rainfall, or in locally wet places, always produces woody growth, either as tongues of woods in river bottoms or the parklike landscape, already mentioned. If, however, the rainfall is too low to produce even the grasses and their associated herbs, an entirely different type of vegetation usurps these still more dry regions, resulting in some of those strange plants of the deserts, among which water storage is practically universal, as is the ability to live for long periods without rainfall.
DESERTS
Of absolute deserts there are none in North America, for no part of it is so dry that plants of some sort do not grow, and in fact in hardly any part of the world are there regions of any considerable extent where plant life of any kind is lacking. There is a small section of northern Chile and adjacent regions on the western slopes of the Andes where nothing grows, and the traveler is met with a cheerless landscape of bare ground and sandy or stony soil. There is no record of it ever having rained in such places, and if there be only a single rain consisting of a fraction of an inch a year, a few plants, usually scraggly low herbs with thick leaves or else quick-flowering annuals will be found.
But there are hundreds of thousands of square miles of the earth’s surface where the rainfall is sufficient to produce plant life, but where it hardly exceeds five or at most six inches in a year, and in some regions is less than two inches. From several different stations in the desert area of our own Southwest the average annual rainfall is about five and a half inches. Such regions produce a desert type of vegetation and are popularly, if not properly, called deserts. The largest is of course the Sahara in Africa, but there are huge tracts in Arabia, China, Thibet, Australia, South Africa, and in some other countries where the conditions for plant life are so unfavorable that only in the better sites are plants found at all. In all deserts there are very large areas entirely without plants, due to shifting sand or other local conditions, which, added to the generally unfavorable climate, make plant life impossible.
With rainfall so low and in most of the regions the temperature so high, the vegetation must be insured against too rapid transpiration. Perhaps the best illustration of how high transpiration both in plants and in man may be in such regions is gained by a statement of D. T. MacDougal, who writes of man traveling in the desert that “the amount (of water) thrown off the skin is correspondingly great, and if the loss is not made good, thirst ensues and ten hours’ lack of water may thicken the tongue so that speech is impossible.”
Under such conditions it is not surprising that desert plants are among the most curious and weird members of the plant world. Every device both to retard transpiration and to store up water to last over a completely rainless period, may be found. In America, to which all the hundreds of different species of cactus are practically confined, we find giant forms, often covered with spines and prickles which prevent their destruction by cattle, and many others that hug the dry sandy soil with curious tortuous branches. None of them have leaves such as plants of the forest or grassland possess, for desert plants cannot afford the luxury of foliage that, because of too rapid water loss, would destroy their chance to survive. Cacti do produce tiny leaves at the ends of their joints, but as if recognizing the inhospitable world into which they are born, practically all of them drop off, so that for the great bulk of the life of most cacti only the bare branches are evident. In most kinds these branches are green, assume the functions of leaves, such as transpiration and the manufacture of food by photosynthesis.
While in cacti and in the giant cactuslike spurges of South Africa the ability to store water is tremendously developed—our giant cereus or saguaro often holding 125 gallons—most desert plants rely upon retarding transpiration for their existence. Leafless shrubs and trees whose often spiny branches are green and perform, on a much-reduced scale, the function of leaves, are among the most common characteristics of desert plants. Some, as Parkinsonia microphylla or paloverde, have tiny leaves which they put out during the spring showers, but quickly lose them as it gets hotter and drier toward midsummer. Many of the plants that do produce leaves regularly have the surface of them so shiny as to appear varnished, or so thickly coated with hairs as to simulate cotton or wool, both of which reduce transpiration. There are many plants, some of which do not even live in a desert but in a locally dry habitat, that also have the utmost development of structure to prevent transpiration. One of the most extraordinary is the vegetable sheep in New Zealand. An inhabitant of dry rocky places, its water supply, although rainfall is fairly abundant, is precarious due to drainage and the failure of the rock to prevent run-off. The different species of Raoulia, of which R. eximia is one of the best known, are admirably adapted to exist under such conditions. L. Cockayne, an authority on the flora of that island, writes of these strange plants: “Perhaps the most striking denizens of rocks are the various kinds of vegetable sheep (species of Raoulia), which form hard cushions, mostly white, but occasionally green—and of enormous size. The raoulia cushions are all constructed on the same plan. Above, the stems branch again and again, and toward their extremities are covered with small woolly leaves, packed as tightly as possible. Finally stems, leaves, and all are pressed into a dense hard convex mass, making, in the case of Raoulia eximia, an excellent and appropriate seat for a tired botanist. Within the plant is a peat made of rotting leaves and branches, which holds water like a sponge, and into which the final branchlets send roots. Thus the plant lives in great measure on its own decay, and the woody main root serves chiefly as an anchor. The vegetable sheep are not inaptly named, for at a distance a shepherd might be misled.” The genus Raoulia belongs to the daisy family, and furnishes another illustration of the remarkable diversity of this largest and probably most recent of the families of flowering plants, which appears to have originated in the Andes and now covers the world.
In Damaraland, Southwest Africa, the most remarkable desert plant was discovered years ago, growing in a sandy and stony plain where the rainfall for fourteen years has not averaged more than two and a quarter inches annually. There are sea fogs, however, upon the condensation of which upon sheets of glass the discoverers of Welwitschia mirabilis relied for some of their scanty water supply. The plant, whose woody stem is deep buried in the ground with only the top appearing above the surface, looks not unlike “the burnt crust of a loaf of bread.” To this only two large leaves are attached. These are many feet long and split into several sections which undulate over the ground very like the tentacles of an octopus. With such strange products of the desert scattered over the plains it is little wonder that Welwitschia caused a sensation only equaled by the discovery of Rafflesia in the rain forest of Sumatra.
While deserts seem to be the most unfavorable places in which plants can exist, and their very existence in many deserts is often a precarious affair, it should be kept in mind that the soil of such places is often by no means sterile. As we found in the section of this book on “How Plants Get Their Food,” water is absolutely necessary for the absorption of food through the root hairs. Where, as in an oasis in the desert, water is locally plentiful a luxuriant vegetation springs up, and one of the most fertile parts of our Southwest was transformed from a desert by irrigation. Then, too, in many deserts there is a pronounced rainy season during which there is a marvelous development of showy flowering herbs that die down as rain ceases or becomes too slight, to wait for another opportunity to make the desert blossom into often gorgeous coloring.
Rain forests, temperate forests, grassland, and deserts—all are immense developments of plant societies depending upon climatic differences for their occurrence. There are some other plant communities which also depend for their development on still other differences of climate. Two such are the vegetation of mountain tops in the tropics, and that strange tundra vegetation near the poles which lives all its life on the ice, only the roots and soil in which it grows thawing out during the brief summer. Temperature rather than rainfall is the cause of these and some other plant societies of more local occurrence.
But what of such well-known plant societies as bogs, in which peat is formed, or the plants growing along the sea beaches all over the world? These, and scores of other plant communities play their part in the distribution of plants, but nearly all of them depend not upon climate, but upon usually purely local conditions of soil. Sandy, actually nearly sterile soil, the acidity of cranberry bogs, the alkaline regions in our own West, the salt lakes and inland seas, regions below sea level, the serpentine outcrops, all the hundred and one differences which local conditions exhibit—all these have a very direct bearing upon plant distribution. It is impossible here to go into the details of the different sorts of plant societies which inhabit such specialized places, nor into the truly wonderful adaptations of certain species to peculiar conditions. But in looking at the vegetation of regions through which one travels it must never be forgotten that its general type, such as forest, or grassland, or desert, or what not, is the result of usually widely operating climatic forces, while many, often quite extensive, plant societies in the region are the result of the local environment. There is often an active struggle as to the dominance of the type dictated by the climate of the place, and the local conditions of soil that tend to nullify general response to it. On Long Island, New York, for instance, there are areas which climatically should produce dense woods of the summer forest type so general all through the Northeastern States. Actually the water-worn sands and gravels that covered the south side of the island in glacial times, are so poor in plant food, that many square miles of this region are now covered only by low scrub oaks and other plants suited to poor soils.
A final word of caution is necessary to those who see in the foregoing brief account of some of the chief causes of plant distribution an answer to questions that many of us ask about why plants or vegetation are of such and such a kind in a particular locality. It has been convenient—nay, it has been necessary—to consider these various factors one by one, but the distribution of almost no individual, and certainly of no widely spread plant community, is the result of any one of these factors operating singly.
The geological history of the region, the links with the past of the species composing the vegetation, the climate, the cooperation of various outside agencies in seed dispersal, the conflict of different species, and of different vegetation types, these and scores of other factors, operating to-day, or having operated in the past—it is all these that are reflected in the plant covering of the earth. The variety and beauty of that covering are too well understood to need further mention here. The extraordinary efforts that the plant world makes to keep all but a minute fraction of the earth clothed with some sort of vegetation we have seen in the pages just turned. No other phase of the study of plant life is so replete with interest as plant distribution. Rightly understood, it is a study, “the cultural, esthetic and practical value of which may well outweigh any other.”
GENERAL CONCLUSION
We have now traced, all too briefly and with the many omissions that such a general account as this makes necessary, the broad outlines of plant life. From the architecture of their outer characteristics, which takes up the first chapter, we have gone step by step into the story of what goes on within the plant, and how it reproduces its kind. These actions or behavior of plants have resulted in many things of great practical importance as well as being of absorbing interest in themselves. What some of these results have been, we see reflected in the uses of plants to man, in the history of their development, and, most of all, in the way they are distributed over the earth to-day.
If plants are still “just plants” to most readers, this book has been written in vain. Those who have gleaned from its pages some conception of what a fundamental thing plant life is, will doubtless want more information than could be included here.
If any considerable proportion of the readers of this volume feel that they have already outgrown it, and that they have many questions about the plant world for which they will have to go to more specialized works for the answer, then this book has more than fulfilled its mission.