CHAPTER VII: HISTORY OF THE PLANT KINGDOM
1. Dawn of Plant Life On Earth
“And the Earth was without form and void”
THE quotation from the second sentence of the first chapter of Genesis tells us more in eight words than could very well be said in as many chapters. Not only have we biblical authority for this early absence of life on the earth, but all the accumulated knowledge of the ages points in the same direction. We have already seen that plants, because they can take inorganic substances from the earth and air and transform them into organic food, must in all probability have come on the earth before animals which, directly or indirectly, all rely upon plants for their food. Even those animals that eat only flesh devour other animals which depend upon plants for food. It may safely be repeated, then, that upon plants all animal life depends, and that, in the dim beginnings of things on the earth, it must have been some form of plants that were the first living things. Extremely simple unicellular animals, however, are known to date from early times.
In the volume devoted to that subject in this series, you will find that at the very earliest stages of what we know as our globe there was a segregation of land and water somewhat different from our great oceans and continents to-day in extent and area, but differing mostly in this—that much of the water was fresh and very nearly boiling hot. We have still the remnants of those great reservoirs of hot water, as our hot springs and shooting geysers only too well prove. And if all plants were as quickly killed by hot or boiling water as the common garden geranium, we should not expect any plant life to have developed upon the earth until all those great bodies of water had cooled. To have waited for that would have been to delay the appearance of plants for no one knows how many millions of years, and there is some fairly good evidence that long before normal conditions of heat and cold were established there already flourished certain kinds of plants. What those plants were is something of a speculation, and indeed exactly what they were no one knows. But in our present hot springs grow certain plants, microscopic in size, but quite obviously related to the algæ or to certain bacterialike organisms. They live with apparent comfort and reproduce themselves freely in water so hot that no other form of life will maintain itself. While there is no proof that these present plant inhabitants of hot springs, common in the West, are descended from infinitely ancient progenitors, it is a fair assumption that some organism capable of growing in warm or hot water was the first living thing to appear in a world otherwise “without form and void.”
This great question of how plants came on the earth, and particularly how from these apparently simplest organisms our whole wonderful vegetation has arisen, has always been one of the most interesting things in the history of the world. There are many different ways of studying this, and in the very earliest stages of plant development we are forced to reason, not so much by actual records or buried skeletons of the plants that probably existed then, for only a very few have ever been found, but by our knowledge of the physical and chemical requirements of unicellular plants, and those slightly more developed, and of their individual life histories. It is, for instance, certain that the first plants must have been aquatic, as no real land plants are known for hundreds of thousands of years after the earth was quite capable of maintaining plant life. The absolute necessity of water to complete fertilization in nearly all cryptogams also makes it fairly certain that water plants, and these of the simplest nonflowering type, were the first living things to be found on the earth. And it is more than a fair inference that these were inhabitants of warm or hot water. Subsequently, as the water cooled, they may well have been not unlike the green scum found on ponds to-day.
Of course, the actual origin of life itself is still as much of a riddle as it was when the ancient philosophers began to speculate about it years before the Christian era. Protoplasm, the unit or basis of all life, while its composition and growth requirements are fairly well known, has never been made in any laboratory. Nor have scientists ever been able to decide what the combination of physical and chemical forces must have been to originate it. But that from a perfectly sterile, probably steaming hot globe, there did finally develop some form of life, and that this must have been aquatic plant rather than animal life, seems not only certain, but the only hypothesis upon which all subsequent development of life must have been based. It is not necessary to ascribe the origin of life to providential inspiration nor to the meddling of strange and outlandish deities, as all savage tribes did and some more civilized peoples still do. There can, however, be no escaping the fact that life is more than the combination of physical and chemical conditions which sustain it, and that its origin has never, and may never be “explained” by merely describing the conditions which unquestionably favored its appearance. In other words, the origin of plant life throws us back upon things believed but incapable of proof, and is none the less wonderful because we cannot yet understand the probable progression of forces and materials to which it owes its origin.
Assuming, then, and we must all start with this proposition, that aquatic plants certainly, and warm or hot water plants probably, were the first living things upon the earth, what are the next steps in the history of the plant kingdom? The answer to that question involves a few simple facts in geology and, particularly, in the making of fossils, which must be understood before we can see those steps or their significance. The geological changes which have resulted in the present condition of the earth’s surface are described in the volume devoted to that subject, and will not be repeated here. But some mention must be made of the formation of fossil plants, particularly as it is upon the evidence of these that the story of the development of plant life is literally written in the rocks.
If a leaf or twig drops into shallow water with a clay or mud bottom, it ultimately sinks, and if then a film of clay or silt is brought down by freshets or what not, it will bury the leaf or twig, of course, filling in every slight depression. If then the buried object were raised so that it dried out and could be split open, we should find a perfect impression of the veins and other outward characters of the leaf etched in the clay. This is often such a perfect process that every detail of the leaf is left in the mud impression, and only the opportunity for this impression to become hardened into rock is needed for us to have a fossil. For these are merely the final hardened rock stages of a process that began as we have indicated, and the thousands of fossils that have been dug out of the earth prove how common the conditions for their formation must have been in certain periods of the world’s history.
But of the untold millions of fossils that have been made most have been destroyed, for the geologists tell us that the earth’s crust has been subjected to much upheaval. Mountain chains thrown up, inland seas formed, great river systems carved out, and tremendous periods of vulcanism or fire action have made the earth’s crust, at different periods, a mighty restless place. And these changes, so slow that often millions of years have elapsed before they were completed, have sometimes been favorable to the making of fossils and sometimes to their destruction. Darwin once wrote about the fossil record that he saw it “as a history of the world imperfectly kept, and written in a changing dialect; of this history we possess the last volume alone relating only to two or three countries. Of this volume, only here and there a short chapter has been preserved; and of each page, only here and there a few lines. Each word of the slowly changing language, more or less different in the successive chapters, may represent the forms of life, which are entombed in our consecutive formations, and which falsely appear to have been abruptly introduced.” And yet it is upon the evidence of this fossil record that most of our knowledge of the history of the plant kingdom is based.
The difficulty of getting any true picture of the beginnings of plant life is great, for those earliest stages of development were unquestionably water-inhabiting plants, whose tissue is mostly too soft and too easily decayed to make fossil impressions of them likely to be preserved. Yet fossil algæ have been found in rock strata so old that no fern or flowering plant had yet made its appearance. It is not too much to picture the world then as peopled only by cryptogamous plants of simple structure, living in the water, and land plants which to-day make up the bulk of our vegetation as not yet developed. Furthermore, there is in these earliest stages no trace of plants with any kind of a vascular system, such as all ferns and flowering plants possess. No stretch of our imagination can readily picture the earth as it in all probability was in that period, with no trees or vines or flowers, the land wholly bare of vegetation, and in the water, along its sterile shores, only unicellular or slightly more developed, wholly nonflowering plants. The conditions supporting such plant life existed for many millions of years, and some geologists have claimed that this period of time exceeded all the subsequent ones combined, so that algæ and some other unicellular plant types are the oldest in the world; and they still exist in enormous numbers.
Much later than this, fossil algæ of comparatively complex structure have been found, showing by their frequency and more highly developed characters a more advanced stage in the development of the plant kingdom. So common were these various types of what we now call seaweeds, although most of them apparently lived in fresh water, and so widespread was their occurrence, that this Pre-Cambrian period has often been called the reign of algæ. As yet no other plants had been developed and none of these ancient types had invaded the land, which for millions of years more must have been entirely without vegetation.
2. The Development of Land Plants
After the reign of algæ and other cryptogamous water plants, our knowledge of which is so unsatisfactory because of the incompleteness of the fossil record, there appeared the first evidences of plants that were able to live with “one foot on the land and the other in the water,” so to speak. How many transitional stages there may have been, and what relation any of these may bear to existing plants, is not known, or is, at any rate, so little understood that it is a disputed point. But somewhere about this period there did appear plants capable of living at least part of their life on the land, and possessing in their vascular system a structure of enormous advantage over their predecessors. It is pure speculation as to what this first land plant was derived from, or from what particular group of water-inhabiting plants it took some of its characteristics. Its appearance, in any case, was a dramatic event of the first importance. Not the least interesting feature of it is that the very plants of which we have indisputable evidences of being the first land plants have come down through the ages to the present day. For it is practically impossible to separate our modern representative from its ancient ancestor, despite all the tremendous changes that have been going on both in the history of the earth’s crust and in the vicissitudes of the vegetation in meeting those changes.
There is the best of evidence that these first land plants were of the club moss family, which are relatives of the ferns. One of them, representing our present Lycopodium Selago (Figure 107) so closely as to be practically indistinguishable, is a common type, as revealed to us in the fossil record. The present plant inhabits rich, moist, and mostly evergreen woods in the northern part of the globe, and is common in the Adirondack and White Mountains.
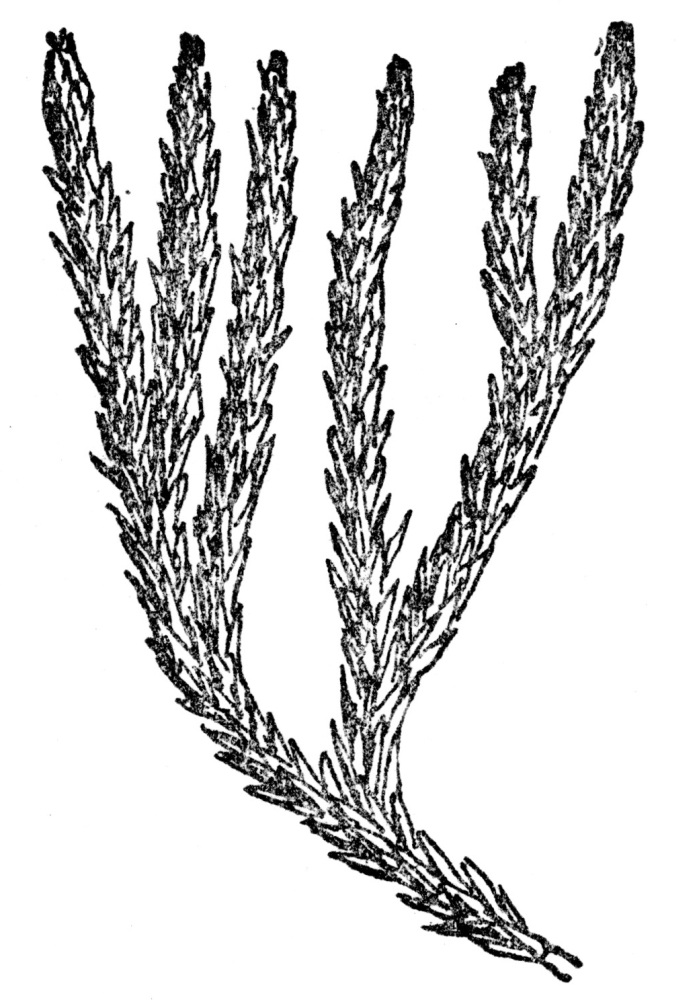 FIG. 107.—CLUB MOSS (Lycopodium Selago)
FIG. 107.—CLUB MOSS (Lycopodium Selago)
A club moss which has come down through the ages almost unchanged from the days when coal was being formed. Grows to-day in the north temperate zone, particularly in mountains.
Nor was the earth peopled wholly by this ancestor of Lycopodium Selago, for we find at this time, or just after it, a great development of plants of this type. Some of these were giant, treelike club-mosses that have been so well preserved as fossils that even their internal structure and spore-bearing characters are well known. Many other strange relatives of our modern club mosses flourished in those days, some of which have wholly disappeared, as have all the treelike forms. These highly organized club mosses, quite unlike any modern representative of the family, appear to have been crowded off the earth by other and subsequent types of vegetation, while Lycopodium Selago, and about thirty related species, have persisted to the present day; not precisely in all cases as they were in this dawn of a land flora, but in many cases with modern structure and reproductive processes so close to the ancestral types as to be nearly identical.
Perhaps nothing gives one a better impression of the tremendous time that must have elapsed before the appearance of these ancient club mosses than the very slight modifications from their ancient condition which their structure at the present time exhibits. While nothing is certainly known of their origin, when they first appeared they were plants with a well-developed vascular system, having stems and leaves quite unlike any of their predecessors’, and a reproductive process almost precisely like their modern descendants. In other words, if they have changed so slightly in all the millions of years since our rock-written records of them first occur, what an infinitely greater period must have elapsed down the dim vista of the ages before their appearance. Of this period, with the exception of fossil algæ, we know practically nothing, and, worst of all, the actual transition from a wholly water-inhabiting flora to these certainly land-inhabiting club mosses may never be known. For, added to the difficulty of water plants being preserved as fossils, already mentioned, is the fact that as they are the oldest, they are found in the deepest strata and, consequently, the hardest to find; and due to changes in the earth’s crust, these ancient fossil-bearing strata have often been much disturbed.
The conclusion appears to be indicated that the origin of a land flora came about with the appearance of these ancient club mosses, which are not mosses in our present-day interpretation of those plants, and that at about the same period many other plants also were found, the whole vegetation resembling nothing that exists at the present time, but many of the different kinds of this ancient flora showed unmistakable evidence of being the progenitors of many plants that exist to-day. What these were, and particularly what they accomplished, both in the history of the plant kingdom and in making the world habitable for man who did not come for millions of years after they were preparing the way, will be considered in
3. Carboniferous Plants and the Formation of Coal
The carboniferous time, or the period when the earth was covered with huge forests of strange shrubs and trees, most of which were unlike their modern successors, apparently had a climate so nearly uniform and seasonless that fossil remains of these plants have been found throughout the world. Even in the Arctic the rock strata show the flourishing of forests that must have needed a climate very different from the frigid condition there to-day, and furnishing indisputable evidence of a warm, most probably frostless, climate practically throughout the world.
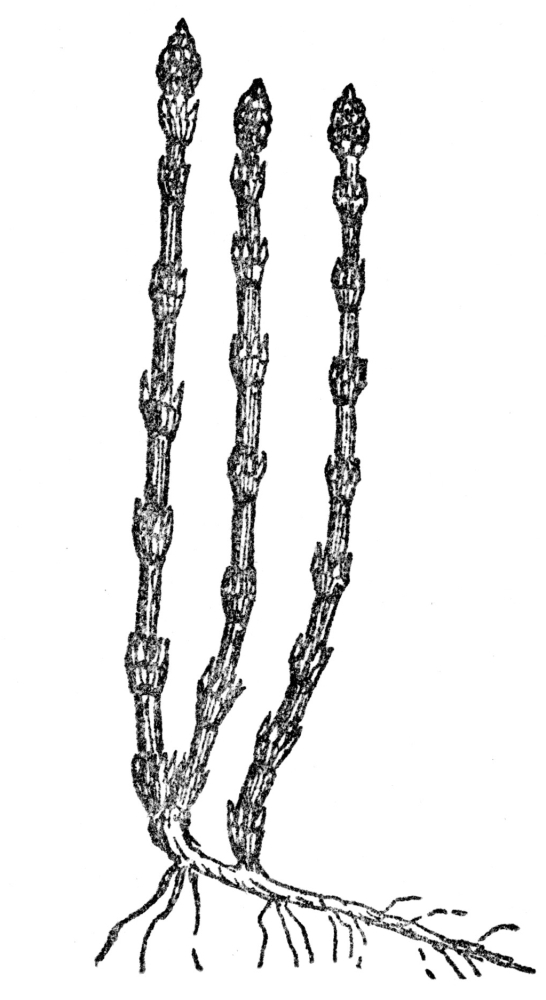 FIG. 108.—HORSETAIL (Equisetum hyemale)
FIG. 108.—HORSETAIL (Equisetum hyemale)
A modern horsetail or scouring rush, common in the north temperate zone. Ancestors of these formed huge forests at the time that coal was being formed.
The giant club mosses have already been mentioned, with their persistence to the present day in much reduced number, and vastly reduced sizes. No one can picture the grandeur of those ancient forests, peopled with queer animals long since extinct and with dragon flies known to have a wing-spread of two feet or more. But with the club mosses were giant horsetails, which in somewhat changed form have also come down to our times, but in much reduced stature and frequently are familiar enough as weedy plants along railway embankments, and sometimes in more natural environments. Most of our modern representatives of the genus Equisetum (Figure 108), or horsetails, are low herbs, but one South American kind still retains the ancient habit of growing to considerable heights, as specimens up to twenty feet high are known. Related to the ancient treelike horsetails were queer vines with slender twining stems, which, judging from their fossil remains, must have been very common. Both the ancestors of our club mosses and the horsetails must have occupied vast swampy areas, as their stem structure indicates a fondness for water, to which, as we have already seen, their still more ancient ancestors were always confined.
Vigorously competing with these plants for occupancy of those great swamps were vast quantities of plants that have been called cycadlike ferns from their likeness to ferns on the one hand and plants like the so-called sago palm on the other. The sago palm, or Cycas revoluta, is a modern representative of these ancient forms, and retains the remarkable characteristic of having its male fertilizing cell capable of movement as we know to be the case in nearly all cryptogamous plants. Yet Cycas, with its related genera, which are found in nearly all the warmer parts of the earth to-day, are true flowering plants which bear cones. We see, therefore, in these old cycadlike ferns one of the first, almost experimental, evidences of the seed habit, and consequently the breaking away from the spore habit which overwhelmingly characterized the reproductive processes of its ancient associates.
The inhabitants of higher parts of that dim, mysterious world, of which we know only that part revealed in the fossil record, were largely ancestors of our modern cone-bearing evergreens. They are known as Cordaitales and have long since disappeared. Forming forests of huge size and making long, slender trunks with a crown of leaves at the top not unlike some modern conifers to which they are, of course, related, these progenitors of our pines and spruces must have been striking objects of that strange landscape. Rooted stumps of these ancient trees have been uncovered, and their narrow leaves, often three feet long, are common as fossils. By some our present conifers and the Cordaitales are both thought to be descendants from a still more ancient group, of which the existence is only conjectured.
We can, perhaps, best summarize our sketch of the plant life existing at the close of this period by saying that all the forms show unmistakable evidence of being crytogamous so far as their reproductive processes are concerned, or else, as in the progenitors of our conifers and cycads, the beginnings of a definite seed habit are indicated. Most of the lowland representatives of this flora were cryptogamous in their characters and ancestry, while some of them, and nearly all the flora of drier sites, appear to have shown the beginnings of flower production. Some of these flowers, which are always cones, are unmistakable as such, and pollen in tremendous quantities has been found among their buried remains. These cones belong to trees that are actual gymnosperms or obvious ancestors of them, for no herbs are known as yet. Nor are angiospermous flowering plants known from this period, nor have any ever been found in strata millions of years younger than the fossil-bearing strata of this age of the ancestors of our modern ferns, conifers, or cycads.
Nor must we picture the development of these different plant inhabitants of that time as passing from one to another in orderly sequence, for that would give us the impression of a regular progression from simple to complex, which may or may not be the truth. There appears to be such a sequence, and the internal structure of the remains of many of these ancient types of plant life have greatly aided our ability to understand their relationships. But with the possibility of various reproductive processes or other structures appearing in quite unrelated forms, and with the comparative paucity of the fossil record in mind, no one can say for certain what are the true lines of descent. The necessity for water in the reproductive act of nearly all the crytogams, the origin of the vascular structure, and the consequent ability to live upon the land, and finally the production of a conelike flower structure with pollen, and all that that implies, are all found during this period.
To the vegetation inhabiting the swamps during this period, man owes a debt perhaps as great as to our modern food plants, for it is upon this, and some later plant remains, that we rely for coal. This period has been well called the Carboniferous, for its chief claim to attention, outside the realms of botanical research, was the deposition of those great collections of plant remains, which, as coal, contain as high as 90 per cent of carbon and furnish the fuel of the world. This is scarcely the place or time to go into the composition of different kinds of coal, but some mention of the conditions under which these ancient swampy forests were transformed into that valuable substance may well conclude the account of a vegetation period the history of which has in large part been found written in the very strata from which coal itself is derived.
In the lowest and wettest parts of those forests there occurred, just as there may occur to-day, a large accumulation of fallen trees and other vegetable refuse. In the ordinary way these would simply rot, due to the work of insects and the fungi of decay, and in a few score years there would be nothing to show. “Dust to dust” would be, and is, the history of so many living things that it is only some machinery for arresting this process which will give us very different results. In the case of coal formation the original impetus appears to be certain microscopic organisms, probably saprophytic fungi, fossil remains of which have been identified, which work upon the fallen mass of vegetation and start its decay, but which can only do so while their prey is still within the influence of the air. The initial stages of decay must, therefore, have been going on while the water was low enough for these organisms to work. But in many parts of that ancient landscape the water level was a fluctuating quantity, due to local conditions or to changes in the earth’s crust. So that many times partially decomposed vegetation masses would become submerged, stopping the work of these organisms of decay that demand air, but providing the only conditions under which certain others could complete the transformation. These bacterial organisms that will work only when deprived of air continue the process, but in a different way. For one thing, the lack of air delays decomposition or almost stops it, as witness the resistance of logs under water, some of which are known to be hundreds of years old. And forest stumps off the coast of Cape May, in New Jersey, are in nearly as perfect a state as when first submerged, over 40,000 years ago. In the production of coal these anærobic (living without air) bacteria release oxygen and hydrogen from the partly decayed mass, leaving as a residue a substance known as peat, which is largely carbon. The transformation of peat into coal depends upon requisite pressure of the strata that may be laid down on top of the peat bed, and probably upon chemical changes that go on after such covering strata have been laid down.
The fact that coal is sometimes found only in thin veins, with layers of shale and other material between, tells us that its origin must often have been a precarious affair, where alternate emergence and submergence would permit first the vegetation to develop and then its transformation to peat, followed by the deposition of fine sands or silt covering the bed. Several such cycles occurred, sometimes separated by untold ages of time, or again by much briefer periods. Certain mines, however, contain over 200 feet of solid coal. The length of time necessary for such a vast accumulation, or how many generations of these ancient plants went into their making, is beyond calculation. With the mining of coal running into the hundreds of millions of tons yearly, we get some idea of how great were those Carboniferous forests, and how extensive they were is proved from the widely separated localities in which coal mines are found.
The Carboniferous age of fern, cycad, and conifer ancestors was by no means a quiet, orderly period, as from geological evidence it appears to have been much subject to alternate emergence and submergence of great tracts of land. Compared with what followed, it actually was a period of comparative quietness, however, and it must, in at least most parts of the world, have permitted the slow development of certain of its plant groups to a state of perfection never reached since. This is particularly true of the ancient relatives of our club mosses and horsetails.
Perhaps one of the most obvious questions to ask about these plants is how long ago they lived, and upon the answer to such a question depend many others. What, for instance, is the position of the Carboniferous as compared to what preceded it and came after? How old is the earth and when did life first appear on it? The evidence upon which such questions are answered comes from the estimates of physicists as to the age of the earth; from students of fossil animals and plants; from astronomers, from geologists and other students. A compromise of these different estimates, and one that has consequently been widely accepted, gives the age of the earth, dating from the time of its having a definite crust with land and water masses, as somewhere near a hundred million years. Such figures are beyond our comprehension and consequently mean almost nothing, but the proportion in time of the different stages of the development of plants may be stated with greater certainty. Taking the total age of the earth as 100 per cent, the period when there is no record of life of any sort may be set down as about 45 per cent of the total, the reign of algæ and development of land plants about 8 per cent, the carboniferous or coal-forming plants about 28 per cent, which leaves only 19 per cent from that distant time to the present. And many things happened in this comparatively brief fifth of the plant world’s history, among them the origin of some plants that have come straight down to us, without discoverable change.
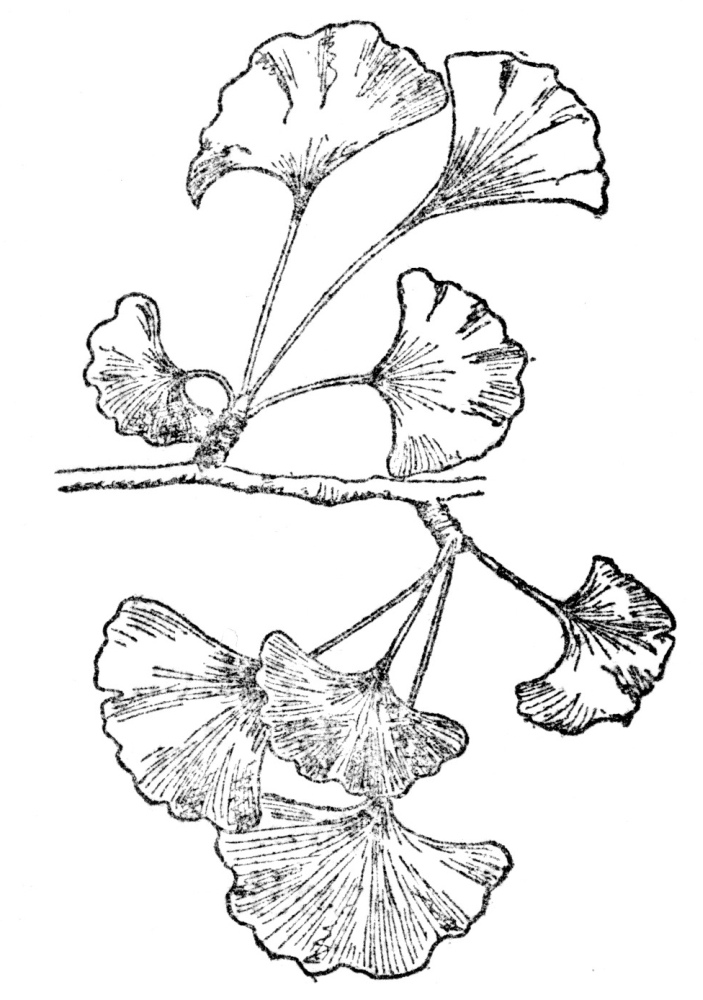 FIG. 109.—GINKGO OR MAIDENHAIR TREE (Ginkgo biloba)
FIG. 109.—GINKGO OR MAIDENHAIR TREE (Ginkgo biloba)
Found in most fossil strata and in a practically unchanged condition from the upper part of the Carboniferous to the most recent fossil records. Now unknown as a wild tree and preserved for us through its cultivation in ancient temple gardens in eastern Asia.
We could hardly leave the Carboniferous time without at least brief mention of the ginkgo tree (Figure 109), or, as some call it, the maidenhair tree. From the upper strata of the Carboniferous it is common, as it is in practically all subsequent fossil accumulations down to the most recent. And yet the tree has never been found wild, although its frequency in temple gardens in China and Japan, always as a cultivated tree, suggests that its disappearance as a wild plant must have come since the priests began preserving it, which can be only a matter of a few thousand years at most. In other words, we have just missed seeing in the ginkgo what has so many times happened to these very ancient types of vegetation, namely, their final extinction. This must have occurred within historic times, and, judging by its frequent use as a temple tree in eastern Asia, that region was its last outpost after its long journey from the dim past. Thousands of other ancient plants have completely disappeared, and one cycad from New South Wales is at this moment putting up a losing fight against modern competitors, but in the ginkgo tree the actual twilight and extinction of its wild existence has missed observations by modern plant geographers by only a brief period. It is almost as though we had waited all our life to see some great event and then missed it by a few moments. Fortunately the tree is now common in cultivation, and not the least interesting feature of it is the fact that its male fertilizing cell retains its power of movement, which dates back to its early associates. Among modern flowering plants only the ginkgo and the relatives of the sago palm or cycads retain this relic of an overwhelmingly cryptogamous ancestry.
The end of the Carboniferous or coal-forming ages was marked by great changes in the earth’s surface, some of them cataclysmic in their effects. What they were in detail is described in the volume on geology and need not be repeated here. What happened to the development of the plant kingdom after this will be considered in the next section of this chapter.
4. More Recent Ancestors of Our Modern Flora
The vegetation at the ending of Carboniferous times was much affected by the great changes in the earth’s surface which happened then. The thrusting up of great mountain chains, the slow encroachment of continental glaciers, and the other phenomena characterizing that period could not but be reflected in the plant population. For one thing the giant club mosses and horsetails were much reduced in extent and finally disappeared, leaving only the immediate ancestors of our present-day forms. Cordaitales gave place to trees not unlike some of the modern yew trees. True ferns as well as the cycadlike ferns with seeds appear to have lived side by side with true cycads, which subsequently supplanted their obvious cycadlike fern ancestors. There was an obvious dwindling of ancient Carboniferous forms, some of which, however, persisted in considerable numbers. Many other plants existed then, some of which died out there, and some of which still survive in descendants, particularly among our conifers and ferns. But there happened toward the upper end of this period an event in the history of the plant kingdom so dramatic, of such far-reaching results, that its appearance might be likened to the overthrow of the Czar in Russian history or to the downfall of the Kaiser in Germany. For with it dawned a new era for the plant world, the effects of which we see all about us to-day.
Somewhere in the rock strata of this period we find the first angiosperm, or plant that matures its seed in a closed ovary, and with the origin of that habit there began such a development of plants of this type that its impetus has not yet been lost. It is impossible to tell at this distance from the origin of that first angiosperm from what it developed, nor how many ages it may have existed before the accident of its preservation as a fossil revealed its presence. It is certainly not without significance that it bore conelike fruits, such as all its associates and predecessors among flowering plants had done, but its possession of large, showy petals is the first evidence of a flower characteristic that was destined to make our present vegetation the lovely thing it is. This exceedingly interesting plant was a Magnolia (Figure 110), or so like our present plants of that genus as to be their obvious ancestor. Somewhere here, too, must have arisen the insect fertilization of flowers which we have seen to be such an important part of flower economy at the present time. Most of this ancient magnolia’s associates must have relied on wind pollination for seed production, as many modern plants still do, but the origin of insect fertilization appears to have come with the appearance of the first really petaliferous flower.
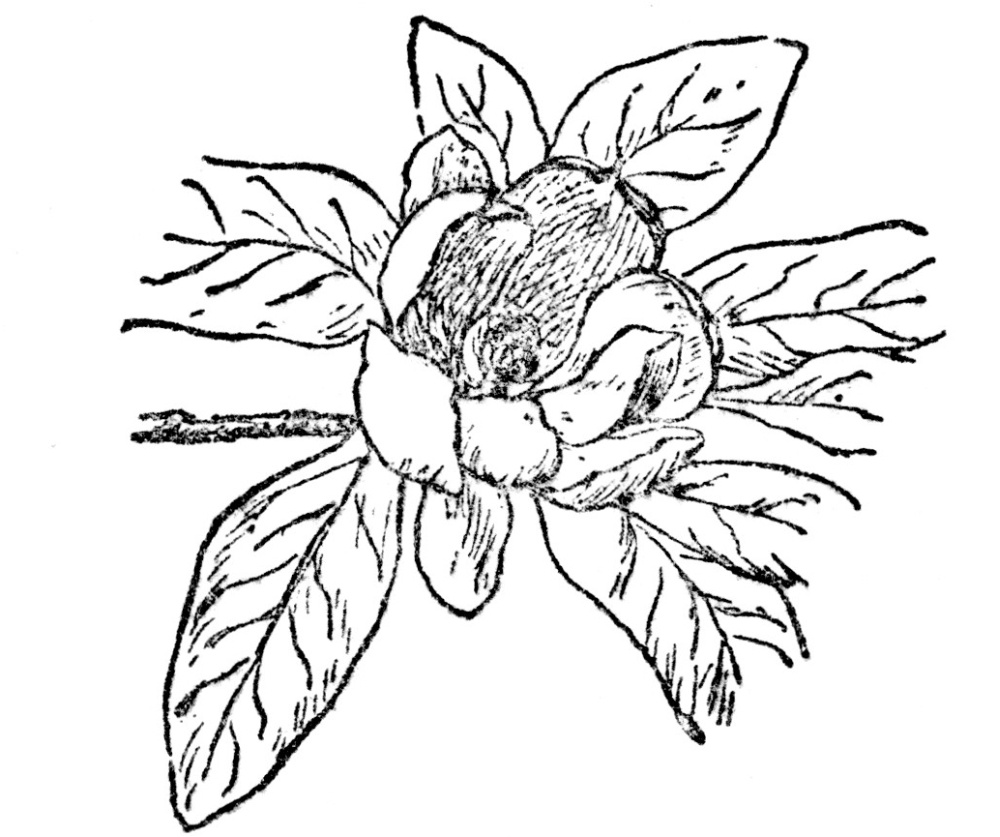 FIG. 110.—COMMON LAUREL MAGNOLIA (Magnolia virginiana) The fossil record tells us that probably the first flowering plant was some ancestor of magnolia.
FIG. 110.—COMMON LAUREL MAGNOLIA (Magnolia virginiana) The fossil record tells us that probably the first flowering plant was some ancestor of magnolia.
With this Magnolia were found other flowering plants which soon increased tremendously in numbers of individuals and differences of structure, such as our sassafras, the tulip tree, the poplar tree, and some others. All of these are trees or shrubs and we do not yet know whether herbs grew in the strange surroundings of that ancient forest or not. Their soft tissue may have prevented their preservation as fossils, but, at any rate, no herb has left its rock-written record from as early in this period as the trees and shrubs. All of these ancient trees have been recorded only in the northern hemisphere and it may be true that this part of the earth was the cradle of all those hosts of the flowering plants that now number over 150,000 species.
There must have been a mighty struggle for occupancy of the really desirable plant sites soon after the rise of these immediate ancestors of our modern plants. For there is every evidence of the progressive dwindling of those still more ancient holdovers from the Carboniferous, and the steady encroachment of the newly arisen and obviously vigorous young race. As we get higher up in the strata, or, in other words, nearer to the present, there are literally thousands of these immediate ancestors of our modern flora, and it is not very long before herbs, particularly grasses and sedges, begin to be common, together with other monocotyledonous plants such as palms. One not unlike our coconut palm has been found in some of these strata in France.
While this period records the origin of hundreds, and there are probably thousands of unrecorded species which are very near our modern descendants of them, it was also a period when the earth’s crust was in an almost constant state of restlessness. Ice periods, huge inland seas, great volcanic upheavals, and the thrusting up of mountain chains such as the Alps, Himalayas, and some others, were only a few of the disturbances to the orderly procession of this wholly new type of vegetation that doomed the older kinds and subsequently conquered the world. The spread of this new element in the plant kingdom was greatly helped and sometimes greatly hindered by land connections between continents, now separated by the oceans. The giant redwoods, now isolated in a few localities on our Pacific Coast, were found then nearly throughout the world. Because of these changes of land areas and some others of even greater influence on plant growth, such as climate, there was a constant shuffling of floral and, of course, animal elements, so that by the end of this period the new type of flora had spread throughout the world, but with here and there very local occurrence of certain genera and families, some of which have persisted to the present day. As we shall see in the last chapter of this book, certain whole families are confined to restricted areas; the cactus and pineapple family, and the genus Helianthus, or sunflowers, for instance, are, with one or two trifling exceptions, wholly American. And we have already seen how many food and other useful plants were first found here by the Spaniards—chocolate, tobacco, corn, the potato, and others.
It would fill the rest of this book to enumerate the plants that flourished toward the end of this period, and, in fact, it might almost be said that the flora of those days was not very different from our own, only it was distributed in different ways and mixed in very different proportions. With the disappearance or partial dwindling of more ancient groups, the rise of the plants that immediately preceded our own ushered in a new era in the history of the plant kingdom.
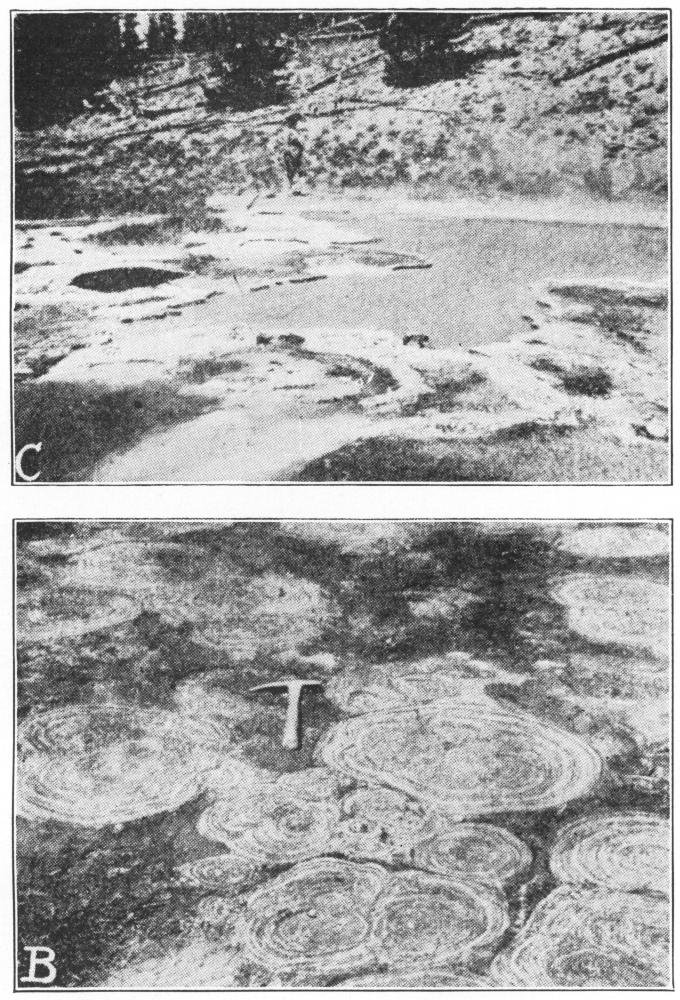
Fossil and Living Algæ Compared. C. A living algal pool colony near the Great Fountain Geyser, Yellowstone Park. (After Walcott.) B. Fossil calcareous algæ. Cryptozoön proliferum Hall, from the Cryptozoön ledge in Lester Park, near Saratoga Springs, N. Y. These algæ, which are among the oldest plants of the earth, grew in cabbage-shaped heads on the bottom of the ancient Cambrian sea and deposited lime in their tissue. The ledge has been planed down by the action of a great glacier which cut the plants across, showing their concentric interior structure. (Photographed by H. P. Cushing. Pictures and explanations of them from “The Origin and Evolution of Life,” by Professor Henry Fairfield Osborn, who kindly permitted their reproduction here.) (Courtesy of Brooklyn Botanic Garden.)

Probable Landscape in the Carboniferous Age. About the Time Coal Was in the Making. (After Patonie) 1. Tree fern 2. Giant ancestors of our horsetails. 3 and 4. Ancestors of our club mosses. 5. Cordaites, a primitive type, or perhaps even the ancestor of our modern evergreens. At this time no herbs and no plants with petals, were known, nor for ages after this period. (Courtesy of Brooklyn Botanic Garden.)
At the end of this period an event of commanding interest occurred, because it happened only some 40,000 years ago. With it came the encroachment from the north and south poles of the last of the great continental glaciers. There had been many before, stretching over a past period of time, but as the last of these great ice invasions it is the most interesting to us. It crowded all these temperate and even subtropical plants that then grew up in the far north toward the equator, and scraped clear of vegetation every part of the earth which it covered. In the volume on geology you will find an account of the extent and thickness of this great ice sheet, which ultimately receded to its present home. As it went back the plants crowded forward to occupy the freshly released land, the far northern or glacial first, followed by waves of other kinds. Some of the glacial or northern plants were left on the tops of the highest mountains, where to-day they persist in complete isolation, nearly all their friends and associates of that greatest of all winters having left them for points farther north. Many students of plant geography think that wave of plant life creeping northward to occupy the region uncovered by the retreating ice is still going on, and recent studies appear to show in at least one isolated mountain in the Adirondacks that the survivals of the ice age which have been isolated on its rocky peak ever since are in considerable danger of being crowded out by invaders from the lowlands.
Not all the geological changes which have remodeled the earth’s surface have been mentioned in this brief history of those plants that preceded our own, nor have anything like all the plants occurring in the different strata been even hinted at. But the thing which has been stressed and for us to fix in our minds is that all our present vegetation literally has its roots deep down in the earth. Some, as Lycopodium Selago, go back no one knows how many millions of years; others, like the flowering herbs, are much more recent. We come to understand how recent we are and what a comparatively brief flash in the pan all our modern development both of plants and in man has been since the last glacial period only by looking for a moment at what has happened in the past. In the account of the Carboniferous plants we found that there remained after that period only about 19 per cent of the earth’s age in which all the changes since then could have come about. If, as may well be possible, this period has been about 19,000,000 years, then the mere 40,000 years since the last Ice Age seems a brief period indeed. As some one has written, to contrast all man’s historic period, back to the days of the most ancient Chinese manuscript, with that long journey from the dim past which the plant world has slowly accomplished, is to realize that we are “as the flashing of a meteor through the sea of night.”
Fossil plants then, and this delving into the dead past of the plant world, reveals to us as nothing else can how much the modern plant kingdom is literally built upon a mighty race of ancestors. Some perished as did the Cordaitales, but left descendants who themselves gave rise to other groups that survive to-day. To look over a list of the fossil plant genera of the different strata is to visualize a drama the like of which no one living will ever see replayed, the results of which are recorded all over the world with its changing panorama of vegetation.
5. Present-Day Plants and Where They Come From
There are living to-day somewhere about 150,000 species of flowering plants; half a hundred conifers or gymnosperms; about 3,500 ferns; 500 club mosses; over 70,000 bacteria, fungi, and lichens, and probably over 20,000 species of algæ. The estimates of the last three divisions are more or less uncertain, as many species are still being discovered.
From what has just been read regarding the plants of earlier periods it is at once clear how completely the flowering plants have conquered all their ancient forbears, and what a pitifully small remnant of once proud and ancient forests are now represented by our club mosses and horsetails. That process of crowding, of the dominance of one kind to the exclusion or even extinction of others is still going on, and, as we shall see in the last chapter, often on a great scale.
If the period just after plants were first known on the earth may be called the Reign of Algæ, and subsequent periods were typified by still other kinds of plants, then our present period is par excellence the reign of flowering plants. In numbers of individuals they are still far outclassed by such cryptogams as the fungi, bacteria, algæ, etc., but their dominating influence in the plant world is unquestioned.
While all our present vegetation must have been derived from preexisting types, all of it is not necessarily directly descended from species which from their fossil records we know to have existed in geological periods older than our own. While the fossil record of the times immediately preceding the last ice age is a much more complete one than for many other periods, it fails to account directly for the great bulk of our varied flora of to-day. While ferns in great variety, gymnosperms, and hundreds of flowering plants are known quite definitely, they total only a minute fraction of these groups to-day. Even granting the always imperfect nature of the fossil record, and we have seen what an accident the formation of a plant fossil may be, and it is common knowledge how few comparatively have ever been recovered—even granting all this, there still remains a large part of our present flora of which the origin probably dates from comparatively recent times. So overwhelmingly true is this that of the Compositæ, or daisy, family, now numbering over 11,000 species, scarcely a handful of fossil species have been found. And in all collections of fossils the woody plants far outnumber the herbs, perhaps because of the greater probability of their being thus preserved rather than to any actual scarcity of herbs in the upper strata. And yet herbs to-day outnumber woody species over two to one. While it is true, then, that our present flora must have been derived from preexisting races, it is also true that much of it is apparently derived from plants that do not date very far back into the past. A few main types of flowering plants unquestionably are to be linked with fossil genera, but these types have now branched out into a wealth of detail that may not have existed and is certainly not recorded in the fossil record.
Some of these types stand out with remarkable clearness, notably magnolia, willows, poplars, walnuts, birches, oaks, figs, sassafras and its relatives, the rosales, the pea family, the spurges, maples, grapes, linden, myrtle, ginseng, and some others. All these, and in not very different aspect from their modern representatives, have been found in the fossils of the different and usually more recent strata before the last Ice Age. But the total fossil record of even these well-known genera is only a fraction of their modern development, and we are constantly confronted with the apparent dilemma of accounting for a present wealth of forms based upon an obvious poverty of ancestry. While the whole race of flowering plants is certainly a new one, as such things are reckoned fossilwise, there has been a fecundity in the origin of new species among these lusty upstarts that is simply amazing. How that, in part at least, has been accomplished will be considered in the final section of this chapter. Not only among these present-day plants, but all through the story of the development of the plant kingdom, we have been reading and writing of the changes of form and structure, some of which have been of far-reaching consequences. It is clear enough that if new types of vegetation and different races of plants have come into being and so modified the complexion of the plant kingdom, those changes must have first arisen in individuals which had within them some capacity for change, and furthermore the ability to use the change to their advantage. While, as we have seen, the losses have been tremendous, no one, with even this brief history of their development in mind, can doubt that there has been progress toward our present perfection of plant life.
6. How Plants Change Their Characters and Become New Species
It was with something very different in mind than the changing of plant characters that Cardinal Newman once said: “To live is to change, and to be perfect is to change often.” And yet nothing better expresses the facts of plants’ ability to change and the results of it than this reply of a great churchman to critics who could not or would not understand the truth of his now famous reply.
It is perhaps best to begin any discussion of the changes in plants by remembering a few simple facts regarding changes in ourselves. “Like father like son” is something more than an old saw which we repeat for centuries without stopping to think whether it is true or only half true. As with so many speeches of the sort, this is just precisely a half truth, for while sons are more apt to be like their fathers than other men, we all have within us the capacity, whether expressed or not, to change very considerably. In other words, all living things may be said to be a reflection or, perhaps better, the result of two divergent tendencies, one of which tends to make like produce like, and the other to produce something different.
Upon the ability of like to produce like rests the continuity of those plant groups, well exemplified by Lycopodium Selago and the ginkgo, which, through all the changing panorama of the history of the plant world, have steadily produced individuals so close to the ancestral type as to be essentially indistinguishable from it. It is upon the possession of this ability that all the different races of plants depend for the unchanged perpetuation of their kind. And, as we shall presently see, it is also upon this very ability that the new forms that do arise, rely for holding fast to their differences.
While it is true, then, that like tends to produce like, it is also and perhaps even more true that they do not precisely do so. In fact, they never do absolutely, and it is the degree of divergence from the type that different plants or animals exhibit, which is the measure of their ability to vary, or “produce something different.” Upon this capacity to vary, from whatever cause, rest all the changes which have occurred in the plant world, and, as we have seen in previous chapters, that has been by no means an insignificant affair. We know, in fact, that while one plant of Lycopodium Selago, than which scarcely any other now living has had greater opportunity to become fixed in its characters, is much like another, no two of them are actually identical. Nor are any two plants of the same species ever precisely alike, any more than two children, even of the same parents, are. The tendency for like to produce like is matched then, or sometimes exceeded, by an almost equally strong tendency to vary.
Heredity on the one hand and variation on the other are the two forces upon which the origin of new species or kinds of plants is based. Both of these work in rather definite ways, some of which are fairly well understood, but many of which are still among the things that scientists are striving to clear up. As the capacity to inherit characters from one generation to another reflects itself in the generally stable conditions which the plant world exhibits, while the capacity to vary is the only source of new forms, it is quite naturally the variations of plants from one generation to another which have been most studied. And the study of variation in plants is not the simple thing we might assume it to be, having in mind only the well-known fact that no two organisms are exactly alike. Wherein do they differ? Are their perhaps temporary or even quite casual differences passed on to their progeny? These and many other questions about variation make it at once the most complicated and often one of the most fruitful subjects of plant research. It is clear enough that with the bewildering variety of different plants in nature it is next to impossible not only to record accurately the amount of variation or its probability of being handed on, but least of all to arrive at any clue as to the origin of that variation. Because of this, and still more because practically no one has ever seen the actual origin of a new species in nature, for we only see the finished product, practically all our knowledge of the laws of variation has been derived from studying cultivated plants. The ease of controlling them and of recording thousands of observations of their characteristics has made the work of the plant breeder, and others who study variation in the vegetable world, much more of an exact science to-day than the mass of often interesting but usually unrelated data that crowd the pages of older botanical literature.
One of the main facts about variation is that it is itself a very variable thing, and the nature of those “fluctuating variations” which are so common in nature well illustrate the point. Within what we know as a species there are many individuals that vary one way or the other from a fairly central, we might almost call them normal, mass of individuals which are typical of that species. Nearly all these forms on the fringe of the species due to the environmental changes and not to changes of hereditary constitution will, if left to themselves, tend through their progeny to become more like the central mass as time goes on, while their position, or some other equally nontypical edge of the fringe, will be taken up by other variants from the average conditions. The amount of this fluctuating variability among plants is beyond calculation, and its action has often been likened to the swinging of a pendulum, which of course spends twice the time passing through the center of its arc, that it does on the limits of it. This very nearly expresses the proportion of fluctuating variants to the mass of typical individuals in many species of plants. In many others, often peculiarly unstable species, the number of individuals at the fringe is very large indeed. Sometimes there may be one or more that do hold their characteristics, in which case we know that they are not true environmental variations but have actually a different constitution. These will be considered presently under another and different sort of variability. But, speaking generally, these fluctuating or environmental variants are merely forms of the species, and, other things being equal, they will not actually originate new species.
It should be emphasized here, and before we go further in our discussion of variation, that species and varieties are after all largely creations of the mind of man rather than the reflection of actuality in nature. When we speak of a “species” it is merely a term which through usage by botanists becomes the symbol of a group of plants more like one another than like anything else. It is obvious that it is therefore necessarily an inaccurate designation of the actual conditions found among plants, which might almost be considered as all belonging to one great group of which, for our convenience in referring to them, we mark off units (families, genera, species, etc.), much as units are marked off on a rule. Species, then, and varieties of plants, notwithstanding the utmost refinement of method used in designating and describing them, and this is historically the most ancient and the most widely developed phase of botanical science, cannot reflect the true conditions, and for a number of reasons. The chief one is that species differ in usually several characters one from another and in large genera there is often a bewildering recombination of characters of the genus in the species belonging to it. Species and varieties are concepts of convenience, nay of absolute necessity, in talking or writing about plants, but hardly expressions of exact truth.
With this in mind we can appreciate the position of those plant breeders who insist that the basis of differences in all plants are the simplest, so-called, factor expressions, which can be isolated and studied with some approach to exactitude in experimental cultures. A factor may be defined as the hereditary determiner or base, which, either singly or in conjunction with other factors, is expressed as a character, such as tallness in peas, or brown eyes in human beings. Such studies have built up a body of information about variation in plants that show it to be of several kinds and with different chances of being passed on from one generation to another.
It was noted in the paragraph before the last that fluctuating variants were sometimes so far off the usual that they might almost be considered distinct forms or varieties. Many such changes appear to be the result of different conditions of the local environment, due to changed conditions rather than to any internal difference in the constitution of the plant itself. A familiar illustration of environmental variation may be seen in lima beans. In any considerable number of plants one often finds smaller and larger pods, either sparsely or well filled with beans. If the beans from the small-podded, few-seeded variants are planted they will produce, apparently quite indiscriminately, large and small podded progeny, just as there will result a mixed progeny if only beans from the well-filled and large-podded kind are sown. In other words the plant fluctuates about a general average which typifies the usual or mass characteristics of the species. One should not, however, regard all variations of the character in lima beans, or any other plant as environmental or fluctuating ones, for some of them may be due to differences in hereditary constitution. And these could only be determined by breeding tests.
Environmental variations are as frequent as the ever-changing conditions of plant growth may determine, and it is common knowledge that such diversity of the environment and the variants resulting from it are extremely frequent.
A much more fruitful source of new forms of plant life results from natural cross-fertilization, which, as we saw in an earlier chapter, is the nearly universal condition in the plant world. If species and varieties can be distinguished only by factor differences, as the plant breeders no doubt correctly insist, it becomes obvious enough that we have in cross-fertilization to consider not alone the factor differences of the pistillate or female, but also of the staminate or male contribution to the union, and how these are reflected in the progeny. Our knowledge of this has practically all been based on work done on cultivated plants under control conditions and it shows some interesting developments which occur from crossing.
If garden peas with, let us say, reddish-purple flowers are crossed with white-flowered ones, the progeny will not be a mixture of these colors but all reddish-purple. If all danger of subsequent cross-fertilization is excluded this first generation of reddish-purple progeny will themselves produce reddish-purple and white progeny in the ratio of three to one. But the extraordinary part of it is that in the third generation all the white and about one-third of the reddish-purple plants will breed true to color. The balance of the reddish-purple plants, which comprise about two-thirds of the second generation, will, if their seeds are germinated, produce colored as against white-flowered progeny in the three-to-one ratio. In other words, these artificial crosses, made by the plant breeder, and this splitting up of hybrids which has been many times verified, are seen to be very fertile causes of the origin of new forms of plant life, if only the factor and character differences in the ancestry be sufficiently complex. With no two plants precisely alike, with cross-fertilization so nearly universal, and with all characters, not a single character or factor expression, as in control conditions likely to be affected by the cross, it may be seen how fruitful a source of new forms this natural crossing may be. It is, in fact, not surprising that plants vary, but that the force of heredity will hold them into such recognizable categories that the red maple, or white ash, or blue cohosh are, with thousands of other species, after all fairly definite designations without which talking and writing about plants would be all but impossible. Some of our most beautiful garden plants have arisen either as the result of natural crossing, or crosses deliberately made by the plant breeder. The scores of forms of the common garden lilac have mostly come about by such crosses, although many other garden plants have arisen by still another kind of variability.
The effects of crossing which have been so briefly noted were not understood, as indeed the cause of them is still unknown, before 1865, when Gregor Mendel, an Augustinian monk, published the results of his work on peas, which furnished the basis for all subsequent work on this kind of variability. His work was neglected until 1900, when what is now known as Mendel’s law, involving the Mendelian ratio already noted, was rediscovered by three independent workers. It is now practically universally accepted as the way in which natural or induced hybrids transmit their characters.
There remains still another type of variability which has been noticed from very early days, and received the name sport, because quite suddenly, from a crop of otherwise similar specimens, one or a few plants showed marked and permanently transmitted differences from the average condition. Such sudden offshoots, which occur rather frequently in many plants, are known as mutants, the process as mutation. Hugo de Vries, a Dutch botanist of world-wide fame, was the chief modern figure who drew attention to mutants, and explained how they differ from fluctuating variants in that while these tend to revert to the average or mass conditions the mutant, once it appeared, held true to type. A well-known example of mutation is the cabbage, brussels sprouts, cauliflower, and kohl-rabi, all of which are sports or mutants from a weedy seaside plant of the mustard family, native in Europe. Since their appearance, hundreds of years ago, they have held their essential characters. If they had been environmental variants they would in all probability have reverted to their weedy ancestor. Hundreds of sports or mutants have been recognized and isolated, so that many of our most valuable garden plants have arisen through this ability of plants to vary in often sudden and rather startling degree. The gardener and horticulturist, from long observation and a keen sight for valuable novelties, have always known that sports are fruitful sources of new forms of plants, but De Vries first scientifically studied them and worked out the principles by which they apparently react. The cause of them is still unknown.
While the cause of mutants has not yet been revealed, we have already seen that the two remaining kinds of variability are due to changes in the environment, or to crossing. Charles Darwin when he published his “Origin of Species,” than which no other book has so completely revolutionized modern thought, did not state the cause of those variations of which he was our greatest observer. He did state the now universally accepted law of the “Survival of the Fittest” which explains how, once these variations make their appearance, the inexorable conflict of nature would automatically weed out the unfit. We have seen all through the course of this history of the plant kingdom how whole types of vegetation have been overthrown to give way to other types better fitted to survive. That process is going on with just as inexorable results to-day as it has down through the ages. While Darwin never claimed that such a purely selective process could initiate new species, many of his partisans who waged battle for him during the first years of the tremendous opposition his views encountered, did so claim, and probably wrongly. The actual cause of the origin of new species, except those demonstrated to result from new combinations of already known characters, through crossing, cannot be explained through the natural selection of plants or animals which exhibit favorable variations. We see their effects; it is obvious enough, that those of value tend for the survival of plants having such variations, and it was natural enough that older students of the problem should mistake these effects for the cause of them. The process of selective elimination constantly going on does tend to fix certain favorable variations and untold millions of plants have had their day in the past due to their possession of such, and the killing off of their less fortunately provided associates. We speak of this great march from the simplest organism up to our most complex plants and animals as their evolution, but we must never forget that it has gone step by step, by one or the other methods by which we have seen that plants vary, or perchance by some undetected method, and that while the results of it are for all to see, the causes of that infinitely slow and quite often wayward variation are not understood. Upon such a conception our modern plant life is seen to be a development of plants that have gone before, that all existing life is derived from preexisting, and not from providential interposition or special creations. All through the long marches of plant evolution there appears to be a definite and final goal toward which it tends, but we do not know the direction, least of all the object, of that goal. In fact, there may be many goals, just as there are the diversity of ambitions among human beings. In tracing the present ascendancy of our flowering plants from their links with the past perhaps we shall find no better statement of their present condition or destiny than to repeat Cardinal Newman’s reply: “To live is to change, and to be perfect is to change often.”