CHAPTER IV: THE FAMILIES OF FLOWERING PLANTS AND THEIR RELATIONSHIP
THERE are perhaps over 150,000 different kinds of flowering plants known in the world to-day, but the flowerless ones are fewer than these in numbers. No one really knows how many thousands of the cryptogams there may be in the world, for all of them have not yet been described, and there are doubtless thousands of which we merely suspect the existence. Flowering plants are so much better known, and have for 2,000 years been the subject of scientific writings, that their relationships and obvious groupings into families are fairly definite and often easily recognizable.
In our ordinary discussions or gossip of neighbors or relatives, the absolutely necessary starting point is to know their name. Their acquirement of this by christening, or by the adoption of it through the usage of parents, settles for life what they will be called. Plants are also christened, and that ceremony is one of the most important events in our subsequent discussion of them.
Plant Names and How They Are Acquired
As we always have at least two names, one to show that we are a Smith, for instance, and another to fix us as John Smith, so all plants have two names, sometimes three.
And because plants come from all over the world and are studied and loved by people of many different languages, it became necessary very early in the descriptive writings about them to hit upon some device that should insure the name of a particular plant being the same all over the world, whether used by a student in the Imperial University of China, or by a garden enthusiast in Connecticut. At the time when this need for christening plants with names that would pass current throughout the world was getting to be a crying necessity, the language of all learned men was Latin, so it was natural that they should give Latin names to plants. That practice has continued to the present day, and there are even now some botanists that cling to the old custom of describing the newly christened plant in Latin. In the olden days this was always done, so that much of our knowledge of plants has come down to us from early books written wholly in Latin. The unfamiliarity of Latin to most of us, and the terrifyingly difficult spelling of some plant names, has resulted in many people saying: “God made the flowers, but the devil gave them names.” Nevertheless, these Latin names are the only ones we can use without endless confusion, just as we bear the names assigned to us by our parents, and no others.
If we were to go out into the country and pick up a wild rose which seemed to be different from any other rose, it would be necessary, in order to talk about it subsequently, to give it a name. After carefully searching through all the books about roses and finding out that it really is a new kind of rose rather than merely being new to us, we should then be ready to christen our new find. As we have said, all plants bear two names. One of these is a general one, like Smith, for instance, and the other more specific, like John. These general names of plants, and they are always their first names, are, because they fix the plant as belonging to a particular group, known as generic names. The generic name of violets, for instance, is Viola; of buttercups, Ranunculus; of wheat, Triticum; of corn, Zea; and of roses, Rosa. Our new rose then bears, without any act of ours, the generic name Rosa, which was applied to roses many years ago, and must therefore be used for all subsequently discovered roses. This generic name of Rosa, like all other generic names, tells us that roses are a well-recognized group of plants, all more like one another than like blackberries, for instance, and because of this they are said to all belong to the same genus. A genus (plural, genera) is a group of different plants, all more like one another than like anything else. To go back to our new Rosa, we must now apply its second and more specific name. If it were a white rose, and had never before been described, we should almost certainly use for its second name something signifying its color and assign alba as the obvious Latin equivalent of white. The second name is always called the specific name, because it shows us that from all other roses our new Rosa alba differs in being white. It is of the genus Rosa, but it is also and forever after a recognized member of Rosa, to which a specific name has been applied—in other words, Rosa alba is a species of rose. Species are thus plants more like one another than they are like any other member of the genus to which they belong. Rosa alba is a species quite unlike Rosa lucida, or Rosa carolina, or all the other scores of roses already known or described. At the time of christening Rosa alba, we should not only enter its name in a book, as ours would be in a parish register, but do much more than that. We should so carefully describe it and, preferably, illustrate it with a picture that no one coming after could ever mistake Rosa alba for any other rose. It can be readily seen that the christening of new plants is very nearly as serious an affair as christening babies, and furthermore, it is only to be attempted by experts. Because this has not always been done, many plants have been christened two or three times. Of course, these subsequent christenings do not seriously matter, for plants, like ourselves, should have only one specific name, the first applied to them. But their subsequent christenings by the careless and ignorant have enormously increased the difficulty of talking or writing about plants. These spurious names are common throughout the literature of botany and are known as synonyms.
The thing to remember about plants, so far as our need for classifying them is concerned, is that they belong to different species which might almost be considered the unit or simplest recognizable category into which they may be sorted. For convenience, we sort species into genera which may well be considered the next highest category in which plants are grouped. The grouping of genera into tribes, of tribes into families, and of families into still larger categories, has nothing to do with their names, but everything to do with our understanding of how they are related to one another, and what these different categories mean in the great collection of plants all about us. In other words, it reduces to a definite system an apparently hopelessly mixed-up mass of plants that, without some contrivance of the sort, would simply be a lot of totally unrelated specimens of plant life. Actually they are grouped in fairly definite categories, some of which are easily recognizable, and all of which fit into that great scheme of nature where everything may seem chaotic, but to the observant it is really a very pattern of order. What it all means and how plants have been grouped into families will be explained, now that we understand how they have acquired their generic and specific names.
PLANT FAMILIES AND ORDERS
A scientist once visiting in Bulgaria noticed that the peasants in that country frequently lived over a hundred years and, in trying to find out the reason, he discovered that they drank large quantities of sour milk. This is alive with a definite kind of bacterium that is of great benefit to the digestive apparatus, and therefore helps in the prolongation of life. In Bulgaria, in other words, a certain food habit of the people has resulted in a definite prolongation of life and fixes that population as of somewhat different characteristics from people not addicted to sour milk. In Japan a whole race lives largely on fish and rice, and while this is not the cause of their yellow skin, it is almost surely the cause of their generally small stature. Many of the English are tall, light-haired, and blue-eyed people, fond of outdoor life and sports, and among the most highly developed of the peoples of the earth. The climate of that island, their generally large consumption of meat and the outdoor life of so many of them, have resulted in quite definite characteristics that make the typical Englishman an easily distinguishable type.
In studying man we are able not only to divide him into such broad divisions as white, black, and yellow races, but due to their particular country or mode of life there are scores of racial subdivisions of these larger categories that everyone recognizes. Such differences are often based on stature, shape of head, mental characters and many others, but those still finer shades of difference between, for instance, a Connecticut Yankee and a plantation owner in the South, are, while noted by everyone, very difficult to accurately describe.
In attempting to find such major differences in plants, some structural character that would set off one large group of plants from every other group, the botanist has a harder task than the person studying man. For all those differences of language and mentality that make up such a large part of our common knowledge of the different peoples of the earth are characters that are foreign to plants. We are thus thrown back on structure as the chief way in which plants differ, and because their reproductive organs are their most important ones, and therefore least likely to vary, it is upon certain characters of these organs that all flowering plants have been divided.
In the chapter on “How Plants Produce Their Young,” we found that most flowering plants have their ovules in an ovary which, after fertilization, develop into fruit and seed. But some plants, while they have ovules, only bear them naked or between scales, never inclosed in an ovary. This is true in all pines, spruces, hemlocks, and all the host of their generally evergreen relatives. Such trees bear cones, between the scales of which are perfectly naked ovules that develop into seeds (Figure 77) that have never been hidden in an ovary, as have the vast majority of the seeds of other plants (Figure 53). These naked-seeded plants are known as gymnosperms or literally gymnos, naked, and sperma, seed, and comprise all the cone-bearing trees in the world, the larger part of which are always evergreen. In some past ages such trees made up the bulk of vegetation of the earth, but at present they are much reduced in numbers. Familiar examples of these Coniferæ, or cone-bearing trees, are larch, spruce, fir, pine, hemlock, juniper, and yew.
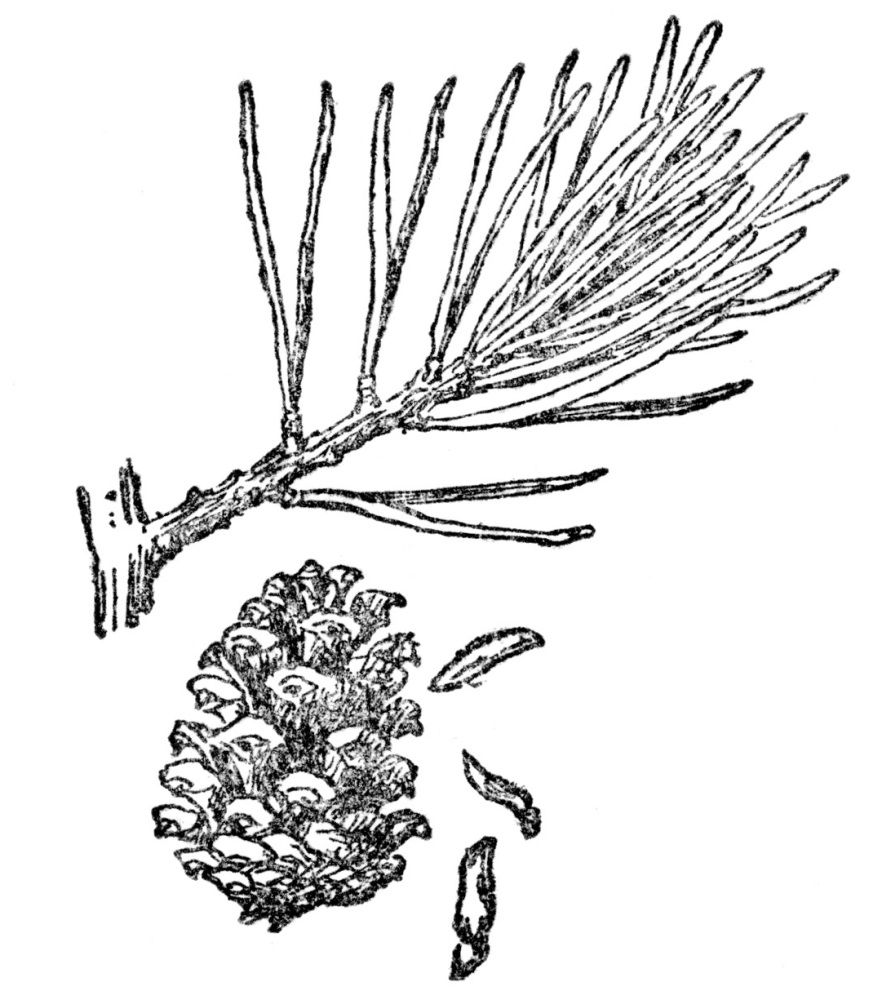 FIG. 77.—THE JERSEY PINE (Pinus virginiana) A gymnosperm or naked-seeded plant. Note the seeds dropping from between the scales of the cone.
FIG. 77.—THE JERSEY PINE (Pinus virginiana) A gymnosperm or naked-seeded plant. Note the seeds dropping from between the scales of the cone.
Most of these are evergreen, which does not, of course, mean that they bear the same leaves always, but that only a few drop off at a time and are so constantly renewed that the tree is actually ever green.
All other flowering plants always bear their ovules in an ovary and, because of this fact, are called angiosperms, literally angeion, a vessel, and sperma, seed. These inclosed seeded plants comprise the great bulk of the vegetation of the earth to-day. So far as the temperate zone is concerned, nearly all of them drop their leaves in the fall, and the trees belonging to the angiosperms are thus said to be deciduous trees.
No better idea of the present size and importance of these two groups of plants can be gained than to state the fact that perhaps not over 500 different kinds of gymnosperms, all of which are trees and shrubs, are known. All the rest of the flowering plants in the world, comprising over 150,000 different kinds of herbs, shrubs, and trees of infinite variety, are angiosperms and therefore bear ovules in an ovary, followed by seeds in or on some sort of a fruit. It would almost seem as though the simplest way to dispose of this great mass of plants would be to sort them into trees, shrubs, and herbs. For all of them belong to one of these types of plant growth, and the ancient students of plants, just before the time of Christ, actually divided all flowering plants into these three classes. This, of course, threw the coniferous trees in with all other kinds and, as we have already seen, they differ from all other kinds in the important character of having naked ovules.
Here, again, in order to get some system out of apparent chaos, we must fall back on some fundamental character. And, again, it is the product of the reproductive process in all this host of angiosperms which furnishes the clue. In the seeds of many of them the young embryo has folded up within it two seed leaves, while in all the rest only one. As we saw in Chapter I, these seeds germinate either with a single seed leaf, like corn (Figure 85), or with two seed leaves, like beans (Figure 81). Every one of these angiosperms belongs to one of these classes or the other, and perhaps more extraordinary still is the fact that in those with one seed leaf there are associated certain leaf and flower characters, while those with two seed leaves are always very different.
In the monocotyledons, or plants with a single seed leaf, the leaves are practically always parallel veined (Figure 83), like corn and grass, and lilies and palms, and hundreds of others. Also, they nearly always have the parts of their flowers in threes (Figure 84). That is, they have three sepals, petals, stamens, and often pistils, or multiples of three. The common trillium or wake-robin, for instance, has three sepals, three petals, six stamens, and three styles. With a few exceptions, and nature seems to delight in producing a few such, all monocotyledons have this parallel-veined leaf character and flower parts in threes or multiples of three.
Plants which send up two seed leaves (Figure 81), on the other hand, bear practically always netted-veined leaves (Figure 79), and the parts of their flowers are nearly always in fours or fives or multiples of these numbers (Figure 80). The well-known wild geranium has five sepals, five petals, ten stamens, and a five-lobed or five-celled ovary. There is some individual variation from this plan, sometimes one organ and sometimes another having more or less than the regular number. But so overwhelmingly true are these distinctions that dicotyledons, or plants with two seed leaves, and monocotyledons,
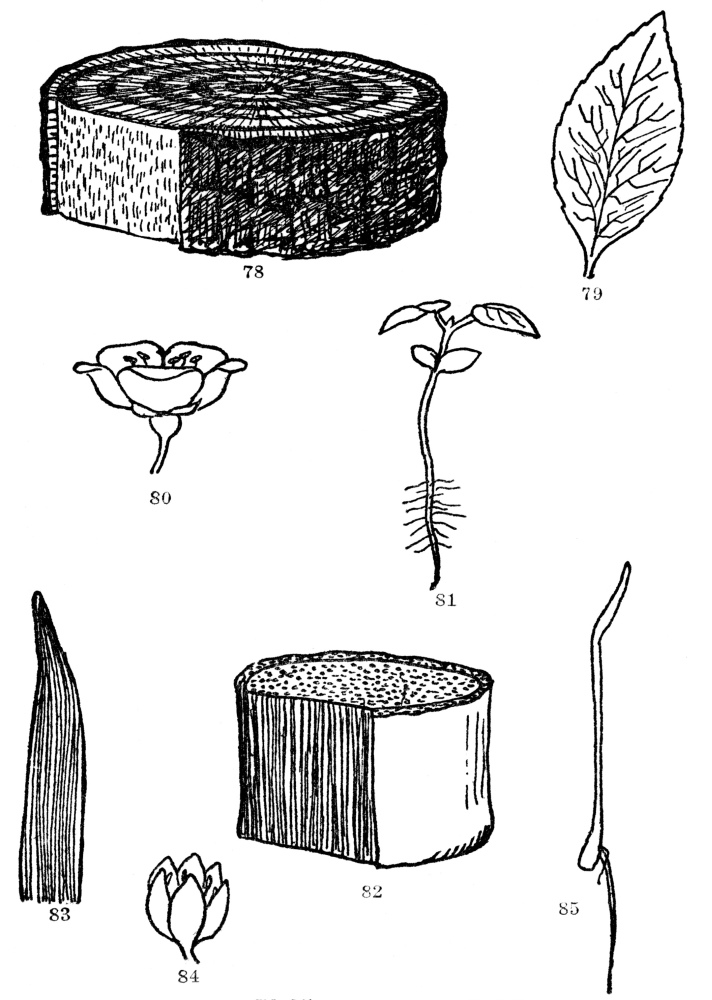 FIG. 78-85.—PLANT FAMILIES
FIG. 78-85.—PLANT FAMILIES
Dicotyledonous and Monocotyledonous growth habits contrasted. Figs. 78-81. The trunk of a dicotyledonous tree showing division of the wood into heartwood, sapwood, and cambium, which the removal of a piece of outer bark exposes. Note the net-veined leaf (79), the seedling with two seed leaves (81), and with the parts of the flower in 5’s (80). Figs. 82-85. Monocotyledonous plant. Note the lack of zones of wood, cambium and corky bark. Such plants have parallel-veined leaves (83), parts of their flowers in 3’s or 6’s (84), and germinate with a single seed leaf (85).
or plants with a single seed leaf, have been for hundreds of years the two great classes into which all angiospermous flowering plants are divided.
Our general view of all the flowering plants may be summarized then as follows:
1. Gymnosperms, or naked seeded plants, include all cone-bearing plants, mostly evergreen and always trees or shrubs. The pine is a familiar example.
2. Angiosperms, or inclosed seeded plants, include all other flowering plants of whatever kind. Divided into: (a) Monocotyledons. Sprouting with one seed leaf, and leaves practically always parallel-veined. Parts of the flower in threes or multiples of three. Familiar examples are corn, grass, sugar-cane, palms, cannas, and lily of the valley (Figures 82-85). (b) Dicotyledons. Sprouting with two seed leaves, and the leaves practically always netted-veined. Parts of the flower in fours or fives or multiples of these numbers. Includes all the remaining flowering plants and is a larger group than the monocotyledons and the cone-bearing plants combined (Figures 78-81).
No matter from what part of the world a totally unfamiliar plant may come, it is always possible to decide into which one of these groups it belongs. That in itself tells us a good deal about its ancestors and its future, “places” it, in fact, in one of those major groups into which all plants are divided. No other characters that plants possess are so important in determining their true position in the scale of plant life as those we have briefly outlined. But merely to sort plants into these large groups does not tell us all we need to know about them. For all plants not only belong to monocotyledons, or dicotyledons, or gymnosperms, but also to smaller divisions of these groups. Just as white men are divided into Englishmen, Frenchmen, etc., so there is the greatest necessity of dividing our large plant groups into smaller and more precise categories.
Some of the chief subdivisions of these large groups have been decided upon the fact that a considerable number of plants in them have some character in common, not found in the remaining plants of the group. Among the monocotyledons, for instance, there is a large class of plants that have tiny flowers between dry, chaffy scales, bear no true petals or sepals, all wind pollinated and are all commonly, though incorrectly, called grasses. These include, strictly speaking, two groups; one, the true grasses in which the stem is mostly hollow and the fruit a grain, while the other, with solid stems and bearing achenes for fruits, are the sedges. The grasses form one family and the sedges another, but while they differ in the characters just mentioned they agree in having flowers of the same general type. Families of plants are thus groups of genera, placed together in the scheme of classification, because they are more like one another than like any other such group. Among the grasses, for instance, are corn, wheat, rice, bamboo, orchard grass, Kentucky blue grass, sugar cane, and hundreds of others, all belonging to different genera, but all those genera grouped into a single family because of their generally similar flowers. Just as the Kentucky blue grass has a generic name (Poa) and a specific one (pratensis), the families of plants must also bear names, usually derived from the generic name of one of the chief genera in it. Because Poa is a large and important genus of the grasses, the family is named after it, with the addition of ceæ. Poaceæ is thus the family name of all the grasses. Among the sedges one of the commonest genera is Cyperus, including many species of the galingale or earth almond. From this genus the sedge family has been named Cyperaceæ (Figure 87). So the rose family is the Rosaceæ, the violet family is Violaceæ, and so on through all the three hundred or more families which contain all the flowering plants so far discovered. Going back for a moment to the Poaceæ and Cyperaceæ, the fact that these two large families are different from each other, but have some characters in common, fixes them as both belonging to one order. Orders are thus groups of one or more plant families, all differing one from another, but obviously related and having some characters in common. The order containing the grasses and sedges is named for one of the families in it with the ending ales. Thus Poales include Poaceæ and Cyperaceæ. Rosales include Rosaceæ and several families.
In other words, individual plants are grouped in species, species into genera, genera into families, and families into orders. These orders are themselves grouped into still larger divisions; there are, for instance, twelve orders comprising all the monocotyledons, and about twenty orders comprising all the dicotyledons. Once we have decided that any plant is a monocotyledon or a dicotyledon, our next step should be as to which order it belongs, then its family, its genus, and finally its species. Needless to say, such studies are necessarily of a technical nature, and while the details of them lie outside the scope of this book, the general plan or scheme of flowering plant classification is as we have outlined it above.
This scheme of plant classification has been developed not only for our convenience in sorting plants into definite categories, but more important still to show, if possible, the relationships, and particularly the development from the simplest to the most complex types of plant life. Thus the monocotyledons begin with the cat-tails, which have mere bristles for calyx and corolla, and lead by infinite gradations to the showy and highly complex orchids, which are considered the climax of the monocotyledonous families. While no general account of the plant families can be attempted here, some of the more interesting in both the monocotyledonous and dicotyledonous groups will be briefly discussed.
Some Monocotyledonous Plant Families
Of the simple plants of this group the Grass Family, or Poaceæ (Figure 86), is the most important, for in it are all our turf grasses, the bamboo and sugar cane, besides scores of others. Over 4,500 species are known, and they inhabit every region of the globe. The steppes of Russia and our Great Plains are predominately grassy; in the wonderful bamboo forests in the tropics are also woody representatives of this family. Certain kinds in the tropics grow as vines, with great hooked spines at the joints, so that nearly every kind of growth-form is to be found in the Poaceæ. All agree in having very small flowers, arranged in tiny spikelets, which are themselves grouped in various ways, although the inflorescence is usually some form of spike, or raceme or panicle. The individual flowers are between chaffy scales, of which several make up each spike. Always the lowest two scales are empty, and the flowers begin in the third from the bottom, or
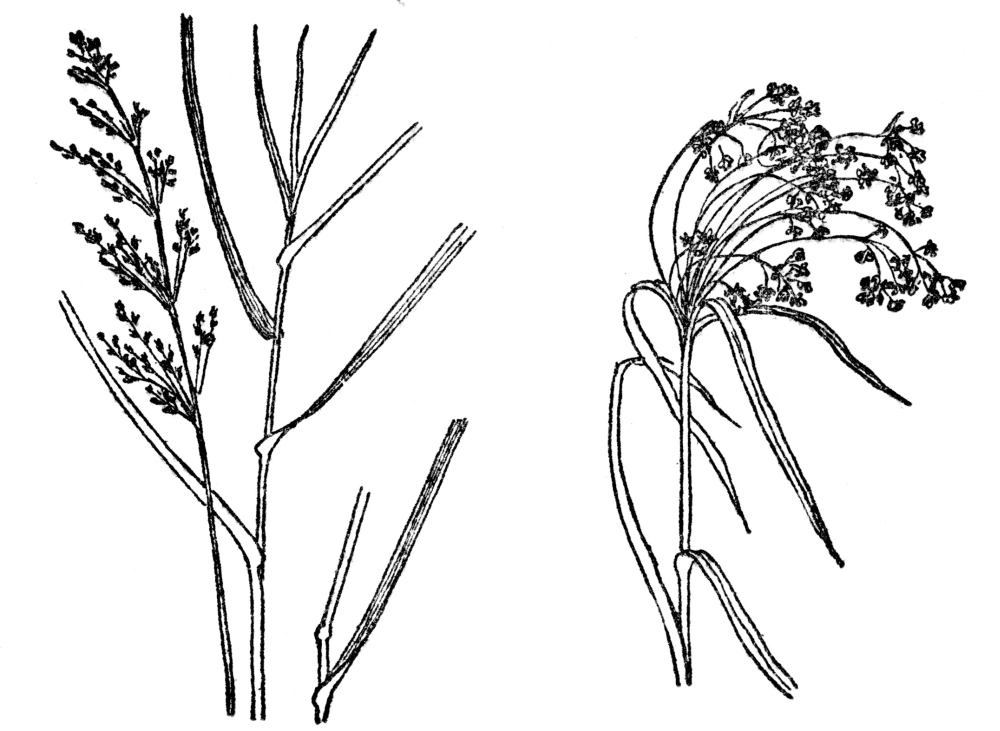 FIG. 86.—BLUE-JOINT GRASS FIG. 87.—WOOL GRASS (Calamagrostis canadensis) (Scirpus Cyperinus)
FIG. 86.—BLUE-JOINT GRASS FIG. 87.—WOOL GRASS (Calamagrostis canadensis) (Scirpus Cyperinus)
Fig. 86. Blue-joint grass, a common grass of North America and a member of the Poaceæ. Fig. 87. Wool-grass, a tall swamp sedge popularly but incorrectly spoken of as grass. It is a member of the Cyperaceæ or Sedge family, which have usually triangular solid stems, whereas grasses have hollow round stems.
sometimes even above that. The flower is so simple that there is neither calyx nor corolla, only three stamens and one to three styles. The fruit is a grain and the Poaceæ, therefore, are the chief source of cereals. Wheat, rice, corn, oats, barley, millet, and rye, all come from grasses, and all, except corn, are natives of the Old World. They were grown for countless ages before the discovery of America, when Europeans first saw corn used by the Indians. As they are wind-pollinated, the flowers of grasses produce no honey nor colored petals, and the vast majority of them have no odor. Most of them reproduce, not only by seeds, but by rootstocks, and many of them grow so closely together that they form turf. In nearly all of them the stem is hollow, and in the largest of them, the bamboo, these hollow stems are used as water and sewer pipes, especially in India. An exception to the hollow stem is the sugar cane, from whose solid stem the juice is pressed out, that is the chief source of sugar; and our common Indian corn.
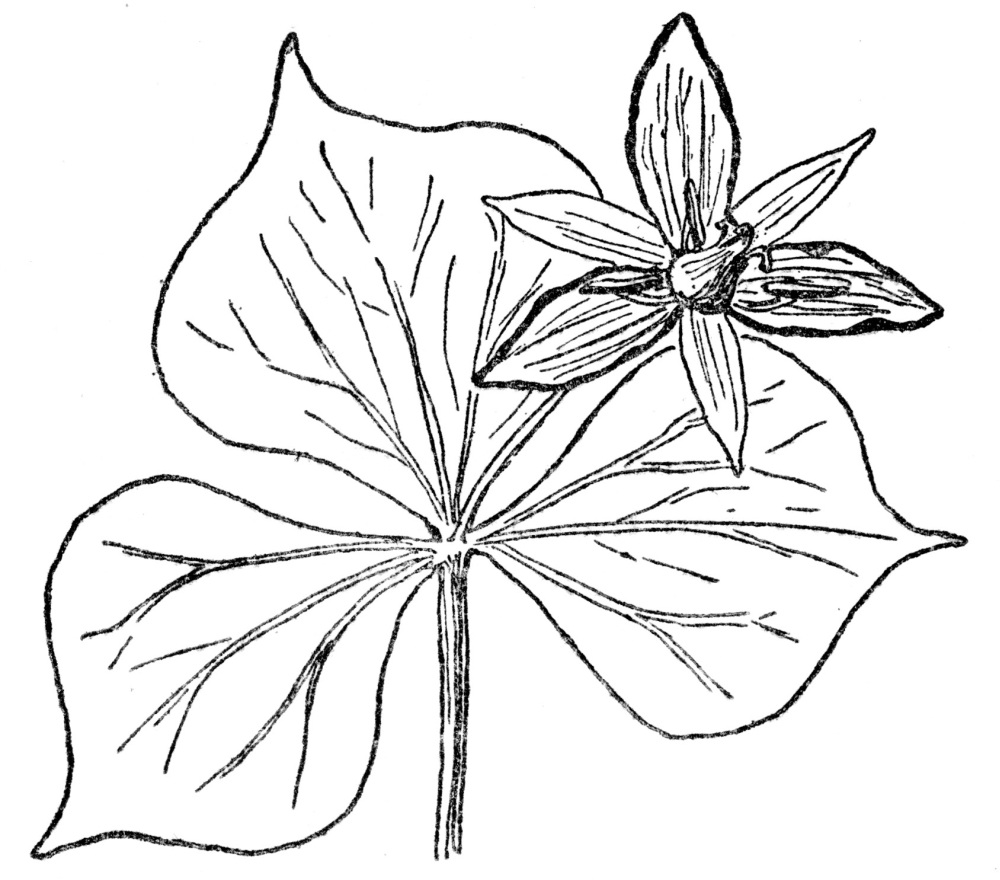 FIG. 88.—SHOWY WAKE-ROBIN (Trillium grandiflorum)
FIG. 88.—SHOWY WAKE-ROBIN (Trillium grandiflorum)
A plant of the lily family (Liliaceæ). Note the tendency to net-veined leaves in a monocotyledonous plant. Such instances are common in nature and net-veined leaves are found in certain species of smilax and most of the plants of the Arum family, containing the jack-in-the-pulpit, both monocotyledons.
Much more highly developed than the grasses is the lily family or Liliaceæ (Figure 88), but comprising less than 1,500 species in about 125 genera. They are nearly always herbs, but the Spanish bayonet forms a woody trunk, while the dragon tree of the Canary Islands is an extraordinary plant for a lily relative, one giant specimen of this being 80 feet tall and over 45 feet in circumference.[1] The flowers in the Liliaceæ are nearly always perfect, that is, stamens and pistils are found in the same flower. Its perianth segments are nearly always six, sometimes distinguishable as petals and sepals, but more often, as in the tulip, all colored similarly. The fruits are practically always a capsule that splits lengthwise. Perhaps the different plants in the Liliaceæ, as well as any others, illustrate the fact that plants of any particular family need not look like one another in order to be included in the same family. Nothing could be farther from resemblance than the bulb-bearing onion, the tulip, the Easter lily, the Spanish bayonet, and the dragon tree. Yet they and hundreds of other plants belong to the Liliaceæ. It cannot be overemphasized that it is flower and fruit characters that determine inclusion in any plant family, and similarity of leaves or habit may or may not accompany such characters. Among other well-known plants in the family, which is found throughout the world, are the crocus, the day lily, the dogtooth violet, hyacinth, and colchicum and aloes used in medicine. Many of them produce bulbs, such as onion, tulip, and lily and some of these contain valuable foods and drugs. The great majority of them are insect fertilized and are therefore wonderfully colored, and some furnish rich stores of honey.
But the most highly developed and interesting of all the monocotyledonous plants are the orchids (Figures 89-92). This family, Orchidaceæ,
 FIG. 89.—ROSE POGONIA FIG. 90.—YELLOW-FRINGED ORCHID (Pogonia ophioglossoides) (Blephariglottis ciliaris)
FIG. 89.—ROSE POGONIA FIG. 90.—YELLOW-FRINGED ORCHID (Pogonia ophioglossoides) (Blephariglottis ciliaris)
Fig. 89. Rose pogonia. A native bog orchid with purplish-pink flowers. Fig. 90. Yellow-fringed orchid. A bog and meadow orchid of the eastern United States.
comprises over 6,000 species and many varieties, the overwhelming proportion of which live in the tropics. Perhaps 90 per cent of them are epiphytes, or air plants, which are perched high up on the branches or bark of trees, and take all their food and water from the air. All the native orchids of temperate North America grow in the ground, however, and their food habits are unique. They depend for food upon a microscopic organism found inside the roots of all orchids, and which helps them to take in the food from the soil. So many of these orchids are partial saprophytes, and without the associated organism they could not grow. Almost uniformly the
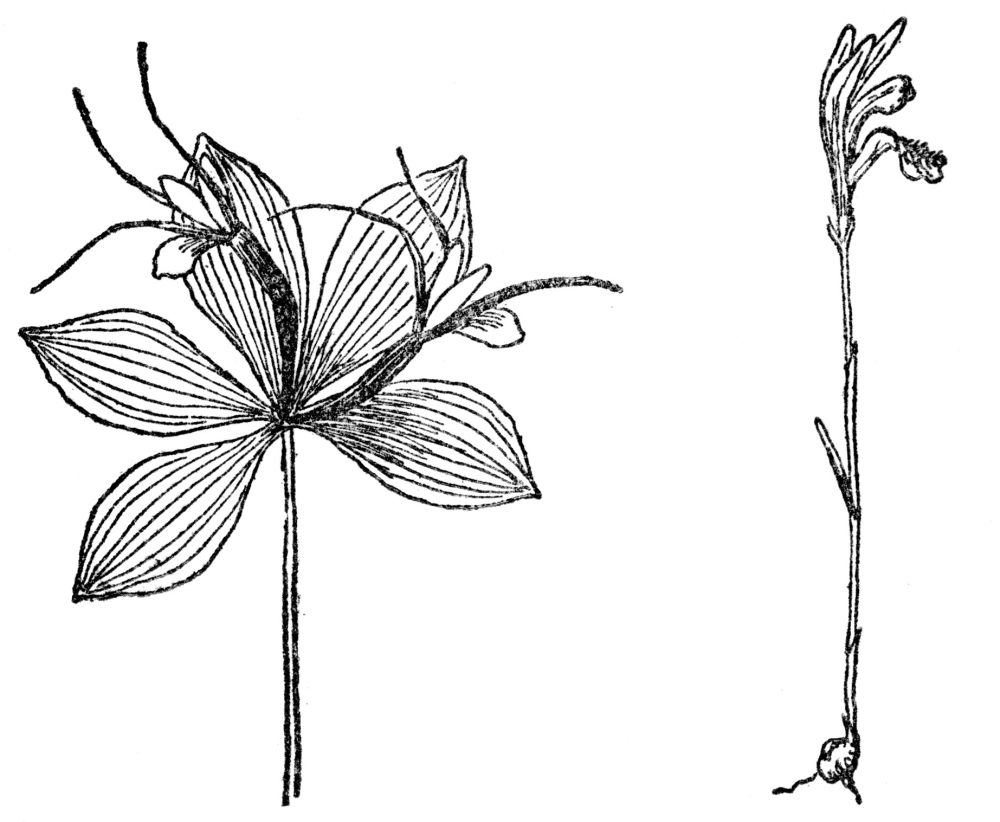 FIG. 91.—WHORLED POGONIA FIG. 92.—ARETHUSA (Arethusa bulbosa) (Isotria verticillata)
FIG. 91.—WHORLED POGONIA FIG. 92.—ARETHUSA (Arethusa bulbosa) (Isotria verticillata)
Fig. 91. Whorled pogonia. A woodland orchid with the leaves and flowers whorled at the apex of the stem. Fig. 92. Arethusa. The most beautiful of our bog orchids, with a fringed lip and pinkish-purple flowers which bloom about Decoration Day. Note the highly irregular flowers in this and Figs. 89-91.
Orchidaceæ have only a very few sheathing leaves, entirely without marginal teeth, and some kinds are practically leafless. The flowers, among the most gorgeous in the world, are always irregular in the sense that there is no obvious series of sepals and petals. Both these are so much transformed as to be nearly unrecognizable as such, but in some orchids there appear to be three sepals. More often of the three inner segments of the flower two are somewhat alike, while the third is quite unlike them and is known as the lip; it is among the most variable of any parts of the orchid flower. As adapted to insect visitors, the flowers of orchids are the most wonderfully developed of all plants. Because of their beauty and strange shapes, orchids have been much sought after by collectors, and explorations of tropical, fever-ridden forests have not infrequently ended in death to orchid hunters. New and rare species of them are constantly being gathered by these collectors. One expedition to New Guinea found over 1,000 kinds never before known, and in the last few decades over 4,000 new orchids have been discovered. For these plants orchid fanciers pay large sums, and a single plant of a rare one sold in London at auction for over $500. The chances of collecting such species made expeditions to the tropics frequent during the latter half of the last century.
These three families, Poaceæ, Liliaceæ, and Orchidaceæ, are perhaps the most important of all the monocotyledons, although commercially the palms, or Palmaceæ, are extensively used. It is impossible to describe or even mention all the monocotyledonous families, but a list of the more important is added. The families are arranged in the order that seems to reflect the development from simpler ones to the most complex, and is the sequence of such families used by nearly all botanists in describing the plant families of the world:
Typhaceæ—The cat-tails. Tall, reedlike swamp plants found throughout the world. One genus and about ten species.
Pandanaceæ—The screw pines. Shrubs or trees with stout, woody trunks and mostly prickly margined, long sword-shaped leaves. Confined to the Old World tropics.
Poaceæ—The grasses. Noted above.
Cyperaceæ—The sedges. Grasslike plants with solid, often triangular stems. Very often inhabitants of wet places. Throughout the world, Crex rugs are made from a species of Carex, the largest genus in the family. About 75 genera and 3,200 species.
Palmaceæ—The palms. All trees or shrubs, or sometimes climbing vines. Includes the coconut and palm-oil trees, two palms of tremendous economic importance. Inhabitants of tropical and warm regions, and only very few found in the United States. Over 130 genera and 1,200 species.
Araceæ—The arums, of which the jack-in-the-pulpit is our best-known native representative. They are nearly always herbs, often of giant size, and the great majority are found in the tropical regions. Flowers very minute, crowded together on a central column (the spadix), and this often surrounded or having at its base a leaflike appendage (the spathe). Calamus root and the skunk cabbage are also native representatives. About 105 genera and over 900 species.
Liliaceæ—The lily and related plants, noted above.
Smilaceæ—Smilax. Mostly prickly vines; our native kinds often called cat briers. Sarsaparilla comes from at least four species of Smilax. Three genera and about 300 species, mostly natives of tropical, but a few of temperate regions.
Amaryllidaceæ—The amaryllis family, noted chiefly for the sisal fiber that comes from a species of Agave, which is one of the many different kinds of century plant. The family has usually capsular fruits and black seeds, and the narcissus and amaryllis of our gardens are well-known members. About 70 genera and 800 species from tropical and warm countries; a few in temperate regions, mostly herbs.
Iridaceæ—The iris, the source of orris root, and containing some our most beautiful garden plants, the blue-eyed grass of fields, and over 50 genera and 100 species are found in this family. Nearly throughout the world, and nearly all herbs.
Musaceæ—The banana and traveler’s-tree. Giant herbs, in the banana having the largest leaves known, frequently twelve feet long and two wide. Natives of tropical and warm regions, and 4 genera and 75 species are known. Flowers often very irregular, and in Strelitzia gorgeous.
Orchidaceæ—The orchids, already noted.
While there are many thousands of plants contained in these monocotyledonous families and in the others not mentioned here, they make up only about one-third of the total number of different kinds of plants known in the world. But in grasses and sedges, in the rushes and a few other families, the number of individuals is greater than in probably any other plant family.
Dicotyledonous Plant Families
All the great bulk of the flowering plants not included in the monocotyledons or the gymnosperms belong to about two hundred plant families that are included in the dicotyledons. In all of them the seed sends up two seed leaves, there are generally netted-veined leaves and the parts of the flower are in fours or fives or multiples of these numbers. In such a large aggregation of plant families there are three well-marked divisions, namely, those that bear no petals or sepals, those that do bear them and where they are separated to form individual sepals or petals, and those where the petals are united to form some sort of a tubular or at least connected corolla. These divisions are perhaps best shown thus:
(a) Apetalæ—Including families where the petals are never present, and in some there is even no calyx. Examples: walnut, hickory, willow, and oaks.
(b) Polypetalæ—Petals present but separate, not forming a tubular or connected corolla. Examples: buttercup, rose, pea, apple, geranium.
(c) Gamopetalæ—Petals united and forming some sort of a tubular or connected corolla. Examples: garden primrose, gentian, salvia, mint, snapdragon, and the daisy family.
Any attempt to describe the families contained in these three divisions of the dicotyledons would take all the rest of this book and crowd out other things about plant life that must not be omitted. All that can be done here is to outline briefly a few families in each division so that we shall have fixed in our minds what the general principles of plant classification are and how these are illustrated by well-known plants. There are many books that deal with this subject in great detail and to them the student should go for further elaboration of the subject. It is one of the most interesting phases of the study of botany, but it demands a longer and more intensive study than can be included here.
(a) Apetalæ—These include families of plants that are the simplest in structure of all the dicotyledons. In all of them there are no petals and in some both petals and sepals are lacking, leaving only essential organs. Taking first those families that have neither petals nor sepals we find that most of them bear their flowers in catkins, a flower cluster familiar enough in the pussy willow. Some of these families are the following:
Juglandaceæ—Trees with compound leaves, flowers in catkins and fruits inclosed in a thick husk. Examples: walnut and hickory. Six genera and over 30 species.
Salicaceæ—Shrubs or trees with simple leaves, flowers in catkins and capsular many-seeded fruits, but no nuts. Containing only willows and poplars. Two genera and over 200 species. (Figure 93.)
Myricaceæ—Shrubs or trees with simple usually fragrant leaves, flowers in catkins and fruits one-seeded. The bayberries. Two genera and about 35 species. (Figure 94.)
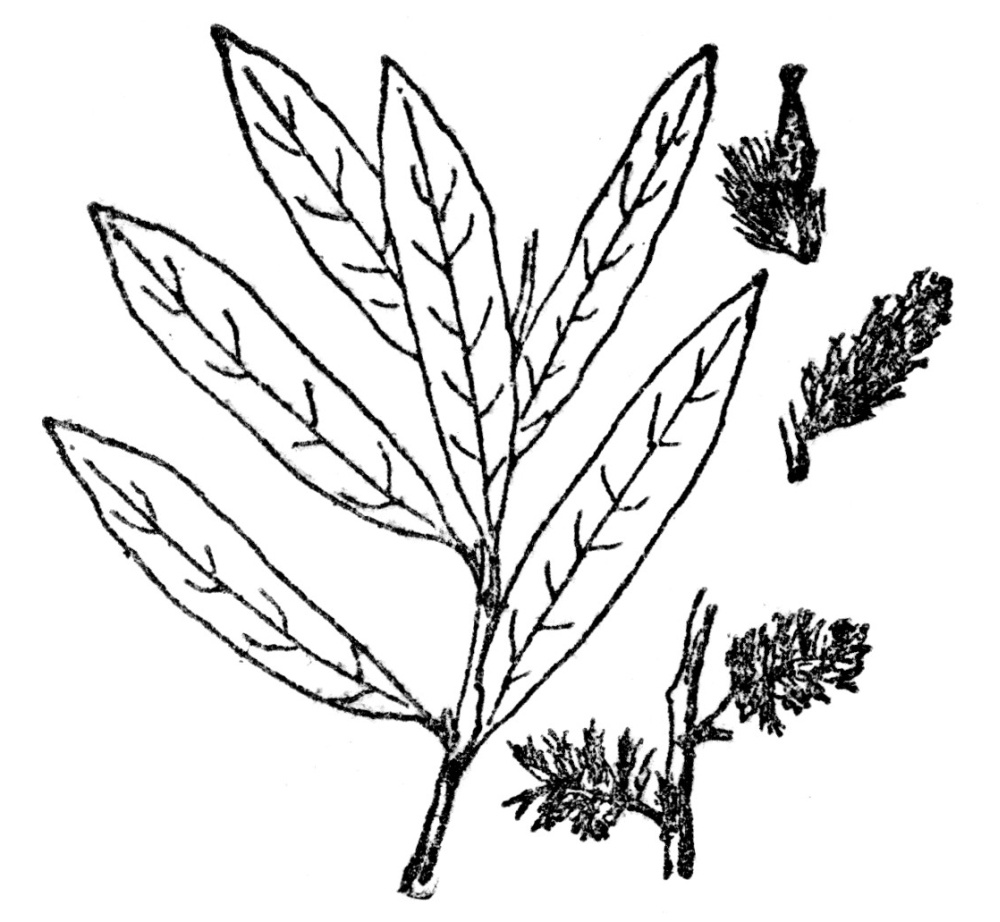 FIG. 93.—PRAIRIE WILLOW (Salix humilis) The Salicaceæ, consisting only of willows and poplars, are always woody plants bearing their flowers in catkins.
FIG. 93.—PRAIRIE WILLOW (Salix humilis) The Salicaceæ, consisting only of willows and poplars, are always woody plants bearing their flowers in catkins.
These and some other families of close relationship are the least developed, in their flower structure, of any of the dicotyledons. All of them bear only the essential organs of reproduction in their very simple flowers. In the walnuts and hickories the different sexes are in different flowers on the same plant, in the willows they are even on different plants, and in Myricaceæ they are often found both ways. All of these flowers are wind-pollinated, so that they bear no honey, usually have no odor, and of course their need for showy petals for attracting insects is nil, although some pollen-eating insects visit them.
 FIG. 94.—SWEET FERN (Comptonia peregrina) belonging to the Myricaceæ. These are usually aromatic, always woody plants, of which several species besides sweet fern grow in the United States.
FIG. 94.—SWEET FERN (Comptonia peregrina) belonging to the Myricaceæ. These are usually aromatic, always woody plants, of which several species besides sweet fern grow in the United States.
Somewhat higher in the scale of plant life are those families that, while still lacking petals, do have sepals. Their flowers are for this reason much better protected against rain or other inclement conditions, which means that they are so much the more likely to reproduce their kind. At least two of these slightly more developed families cling to the habit of bearing some of their flowers in catkins, however. Other families are also found in other parts of the world, but in North America this group is represented by:
 TRANSPLANTING RICE IN JAVA (Courtesy of Brooklyn Botanic Garden.)
TRANSPLANTING RICE IN JAVA (Courtesy of Brooklyn Botanic Garden.)
 A TEA ESTATE IN CEYLON (Courtesy of Brooklyn Botanic Garden.)
A TEA ESTATE IN CEYLON (Courtesy of Brooklyn Botanic Garden.)
Betulaceæ—The birch, alder, hazelnut, and hornbeam. Both staminate and pistillate flowers in catkins. Fruit a small one-seeded nut or a winged samara in the birch. All wind-pollinated. Six genera and about seventy-five species, nearly all from north temperate zone.
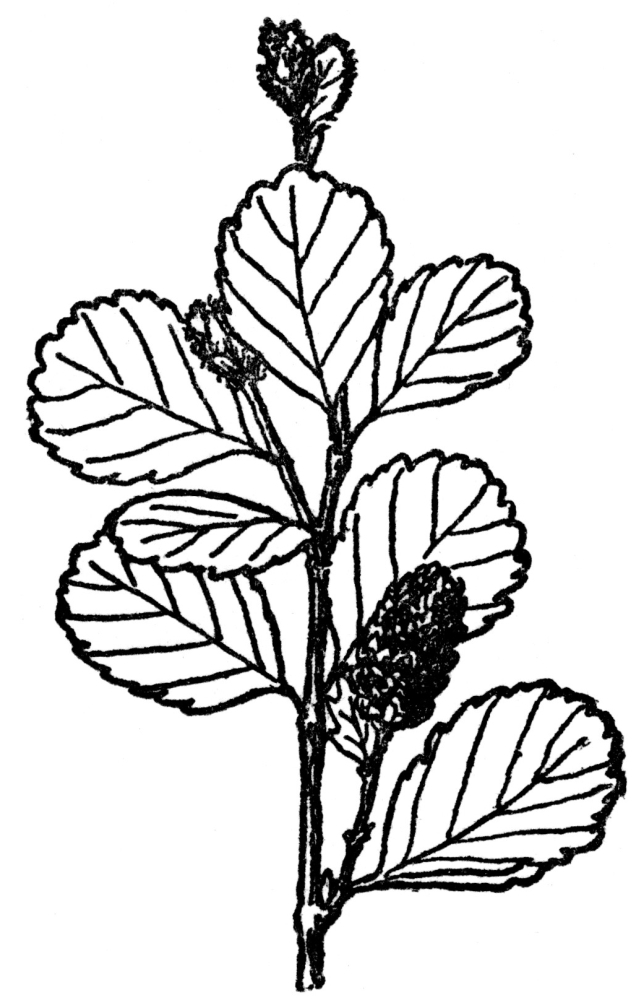 FIG. 95.—LOW BIRCH (Betula pumila) A bog shrub of the Betulaceæ or birch family. Most of them are tall trees with both male and female flowers in catkins.
FIG. 95.—LOW BIRCH (Betula pumila) A bog shrub of the Betulaceæ or birch family. Most of them are tall trees with both male and female flowers in catkins.
Fagaceæ—The oaks, beech, and chestnut. All trees or shrubs with at least their staminate flowers in catkins. Fruit inclosed in a bur (chestnut and beech) or borne in a cup (the acorns of oaks). At least five genera and about 375 species, widely distributed.
The habit of bearing some or all their flowers in catkins which flower usually before the leaves appear, and of having such flowers wholly at the mercy of precarious winds, is, if not lost, at least much less frequent in the remaining families of the apetalæ. All the others, while still without petals, do have sepals and some of these are colored so that insect visitors are likely. There are too many of these families to be enumerated here, but two of the chief are:
Ulmaceæ—The elms and hackberry. Trees or shrubs with minute greenish or yellowish flowers crowded in small clusters or in spikes. Fruit a dry nut or one-seeded and winged; or in the hackberry a drupe, one of the first evidences of even a slightly fleshy fruit in the dicotyledons. About 13 genera and 140 species, widely distributed.
Polygonaceæ—The buckwheat, knotweeds, common dock, and many other genera. Sepals often colored white or pink so the flowers are sometimes at least insect-pollinated. Flowers small and crowded in various clusters, often in a spike. Fruit an achene, a dry fruit familiar enough in the buckwheat. About 40 genera and over 800 species, mostly herbs or vines here, but often trees in the tropics.
From here on plant families leave, with some exceptions, the greenish or otherwise inconspicuous flower color, and somewhere about here they begin to rely more upon insect fertilization for the perpetuation of their kind. None of those so far mentioned have any petals to their flowers, but in the pink family or Caryophyllaceæ we find the first evidences on any considerable scale of the presence of sepals and petals, the latter usually beautifully colored. Familiar representatives of this family are the pink, carnation, chickweed, corn cockle, and the stichwort. There are over 50 genera and 1,000 species, nearly all in temperate regions.
The apetalous families appear to show a development from catkin-bearing trees with the sexes separated, and with neither petals nor sepals, through the Polygonaceæ, with often colored sepals, and the beginnings of insect fertilization. In Caryophyllaceæ, the most highly developed of them all, there are, besides the sepals, often or usually petals, and the reliance on insect fertilization is nearly complete. There are many transitional stages which cannot be included here, but they show step by step the development of the apetalous families from perfectly naked reproductive organs to the next larger group, the polypetalæ, where the process of increasingly complex flower development will now be sketched.
(b) Polypetalæ—In this large group of plant families the petals are free and quite separate, but as if they had not yet lost all the characters of the apetalæ, some families show incompletely the general characteristics of their more stable neighbors. There are, for instance, no petals in many species of the buttercup family, none in the sweet-gum tree nor in the maples, and a few others. But in spite of occasional exceptions this large group of polypetalous families do usually bear separate petals and sepals, and are among the most important of all the plant families. As they number over a hundred and contain thousands of species, all that can be done here is to mention a few typical or important ones. Just as in the apetalæ the families in this large group appear to show definite stages in development from simpler to more complex forms. But the steps are harder to trace and what appears simple characters in some plants are very complex in others.
While all the families in this group have separate petals some of them show a tendency to have united sepals, a character of perhaps some advantage and certainly very common among the still more developed gamopetalæ. Some of the families that have separate sepals agree in having the stamens inserted below the ovary. Of these the following three families may serve as types.
Nymphæaceæ—The water lilies. Aquatic plants with usually large showy flowers in which the calyx, corolla, and stamens often merge one into the other so that it is sometimes difficult to know where one series ends and the other begins. Five genera and 45 species throughout the world.
Ranunculaceæ—Buttercup family. Includes buttercups, clematis, columbine, meadow rue, golden seal, marsh-marigold, hepatica, and scores of other native plants. All herbs, except a few semiwoody vines like clematis. Sepals always present, and where no petals are found, as in marsh-marigold, colored like them. Highly irregular flowers are not uncommon, as in columbine and monkshood. The fruits are berrylike in some genera and in others dry capsules. Thirty-five genera and over 1,000 species throughout the world, but most abundant in temperate regions.
Lauraceæ—Laurel or sassafras family. Includes besides them the guava and cinnamon and camphor trees, all tropical, and the native spice-bush. All trees and shrubs with small, yellow, or greenish-yellow flowers and usually aromatic juice. Fruit a one-seeded drupe or a berry. About 40 genera and over 1,000 species, nearly all tropical, but a few in the United States.
At this point, in the sequence of plant families, there are two or three families that bear quite different fruits than any heretofore noted, and in one of them, at any rate, the four petals are in the form of a cross. So uniformly is this true that the family was for years known as the Cruciferæ, but is better known as Brassicaceæ, from Brassica, the generic name of the mustard. This large mustard family mostly has fruits known as a silique or silicle, which are pods that split into two valves; and yellow or white, rarely pinkish flowers. The juice is always somewhat acrid, familiar through the pleasant pungent taste of water-cress, but none of the family is poisonous. There are over 200 genera and nearly 2,000 species of wide distribution, and common representatives include the cress, mustard, horse-radish, garden stock, sweet alyssum, cabbage, cauliflower, brussels sprouts, radish, and turnip.
Between the Brassicaceæ and the following families there are many others that cannot be mentioned here. Somewhat farther along in the sequence are a group of families, large and important, and all having their stamens inserted around or even above the ovary, and in which the sepals are partly or wholly united. They include some of our most beautiful flowers and useful fruits. Of the many closely related families that agree in these characters the two most important are:
Rosaceæ—Rose family. In the broad sense including, besides the rose, the strawberry, blackberry, apple, pear, peach, plum, besides many herbs with wholly dry fruits. There are always five petals, five lobes to the partly united calyx, but numerous stamens. They may be herbs, shrubs, or trees, with simple or compound leaves, but these are nearly always alternately arranged. There are over 100 genera and nearly 2,000 species. Because of the size of the Rosaceæ and differences in fruit, the apple and its relatives are often included in a separate family, the Malaceæ (Figure 96), and the peaches and plums in Amygdalaceæ. The general structure of the flower is sufficiently uniform, however, for them all to have been included in Rosaceæ (Figure 97).
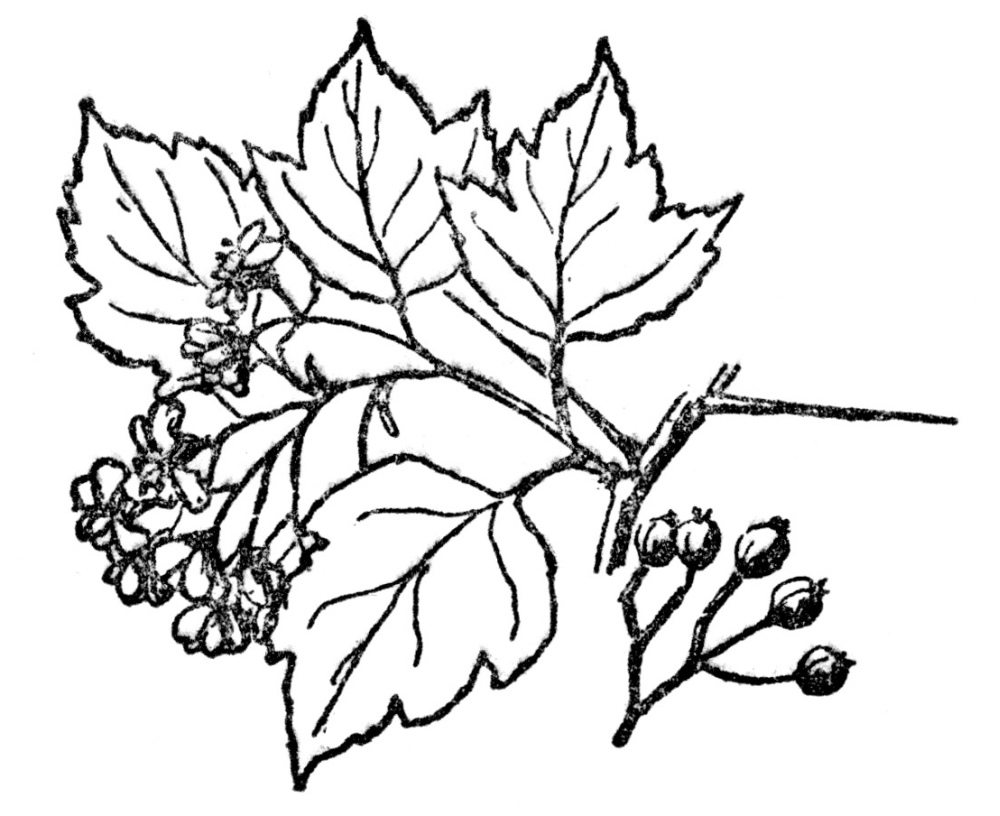 FIG. 96.—WASHINGTON THORN
FIG. 96.—WASHINGTON THORN
A prickly shrub related to the apple, which, with the plums, cherries, pears, strawberry, blackberry and hundreds of other plants are all grouped in the Rosaceæ or rose family.
Papilionaceæ—Pea family. A large family having characteristic pealike flowers, a description of which has already been given in Chapter I. They all bear legumes, a pod that splits down one side, not both, as in Brassicaceæ. Besides the pea, the bean, vetch, alfalfa, lentil, locust tree, and dozens of valuable timber trees in the tropics belong here. Flowers all showy and absolutely dependent upon insects for fertilization. Seeds highly nutritious in many genera, and the roots of nearly all have bacteria associated with them. (See chapter on Food Habits of Plants.) Three hundred and twenty-five genera and over 5,000 species of wide distribution, but most frequent in the tropics.
 FIG. 97.—THIMBLEBERRY (Rubus odoratus) A bristly shrub of the Rose family common in rocky places in eastern North America.
FIG. 97.—THIMBLEBERRY (Rubus odoratus) A bristly shrub of the Rose family common in rocky places in eastern North America.
There are many other smaller families in different parts of the world which hover, as it were, about these two giant plant families that make definite landmarks in the scheme of plant classification. The character of having partly united sepals and numerous stamens inserted around, or even above the ovary, give to all the flowers of the Rosaceæ and related plants a general family resemblance that is very striking. The pea family, and its relatives, also have flower and fruit characters of remarkable uniformity, considering the tremendous number of species.
From this point on to the end of the polypetalæ there are scores of plant families, all agreeing in having a compound ovary, that is, one that is more than one-celled, and in also having quite distinct and separate sepals. Their agreement in these characters, however, ends all other evidences of relationship, and it is beyond the scope of this book to go into the details of each, or even a few of them. A list with some brief notes on those most important must suffice here.
Geraniaceæ—Geranium family. Includes also the common garden as well as the wild geranium. Fruit splitting into five parts. Leaves always divided or even dissected; 12 genera and 470 species, all herbs.
Anacardiaceæ—Includes the tree of heaven, sumac, poison ivy, and in the tropics, the mahogany, all trees, shrubs, or vines; 60 genera and 500 species, mostly in the tropics.
Malvaceæ—Mallow family, including, besides the marshmallow, the rose of Sharon, and cotton. They all have the stamens united into a column or tube which surrounds the style. About 40 genera and 900 species of herbs, shrubs, or trees of wide distribution.
Cactaceæ—Cactus family. Nearly all desert plants, with no leaves or practically none, and greenish stems that function as leaves and also store water. Of the greatest variety of form and always bearing numerous petals and fleshy fruits, of which the prickly pear is familiar enough. Of 40 genera and over 1,000 species, all, but a handful, come from North and South America.
Umbelliferæ (often called Ammiaceæ)—The climax of the polypetalous families, and nearly always bearing flowers in umbels. There are usually many flowers, sometimes several hundred in each cluster. Familiar examples are parsley, celery, parsnip, Queen Anne’s lace, and many others. The seeds often contain an aromatic oil, as in caraway, and some are violently poisonous, as the water hemlock. About 250 genera and over 2,000 species, all herbs, widely distributed, but most common in temperate regions.
We have seen from the foregoing the probable development of dicotyledonous plants from those simplest ones, where, as in the pines, there is merely a naked ovule between scales, through the catkin-bearing trees, without petals or sepals, and all wind-pollinated, to families where just an inconspicuous and, subsequently, a colored calyx is found, and after this the dawn of those plants that have complete and perfect flowers. Among the latter all those so far noted have separate petals, but after the Ammiaceæ, or carrot family, there appears a new character, setting off practically all other dicotyledonous plants from those already treated. This new character—and hints of it are found before it reaches the perfection found in the subsequently described families—is that of the petals being united to form some sort of a connected or, more often, a tubular corolla. The petals are represented merely by the lobes of the corolla, mostly four or five, and in many families of this group, known as the gamopetalæ, literally, united petals, this tubular corolla is irregular and often beautifully formed. In salvia, for instance, there is a hoodlike upper part overhanging the lower tubular part. Other familiar examples of these irregular corollas are the garden snapdragon, Oswego tea, skullcap, pentstemon, and many others.
(c) Gamopetalæ—The earlier families among those generally having united petals seem not yet quite sure of their new character, for a few of them hark back to the condition of having, in some genera, quite separate petals. One of the first families in this series, the Ericaceæ (Figure 98), or heath family, has several genera in which this is true, notably in the Labrador tea and the sand myrtle among native plants, and some foreign relatives. The Ericaceæ are almost exclusively shrubs or trees, but some of our native sorts, such as trailing arbutus and wintergreen, are practically herblike, although they are, strictly speaking, woody plants. The family is remarkable for containing beautiful flowered garden plants, such as the hundreds of species of South African heaths, the heather, the azaleas and rhododendrons, and our beautiful native Rhodora, about which Emerson wrote one of his most beautiful poems. The flowers in the heath family are often perfectly regular and bell-shaped, but sometimes irregular, as in azaleas and several other genera. Nearly all the family rely on microscopic organisms to get their food, and some close relatives, like the Indian pipe, are saprophytes. There are over 70 genera and 1,200 species widely distributed. Central Asia is the home of most rhododendrons and azaleas, scores of species being found in the upper reaches of the Himalayas.
The Ericaceæ are typical of many families in the first group of the gamopetalæ, in that all of them, with a few exceptions like the cranberry, have a superior ovary. That is, the petals and sepals arise from the base or below the ovary, and consequently the mature fruit in such plants is never crowned with the remains of the withered calyx, as blueberries always are and all other gamopetalæ that have an inferior ovary. The character of having an inferior or superior ovary separates the gamopetalæ into two large groups of families, the heath family and many others, with superior ovary, and a few but numerically very important families that always have an inferior ovary.
 FIG 98.—SWAMP AZALEA A shrub of the Ericaceæ, with white or pink sticky flowers and dry capsular fruits.
FIG 98.—SWAMP AZALEA A shrub of the Ericaceæ, with white or pink sticky flowers and dry capsular fruits.
Taking first the families that all have a superior ovary, we must, for lack of space, exclude most of them from here. A few of the most important, or typical, after the Ericaceæ, are:
Primulaceæ—Primrose family. All herbs in which the stamens are as many as the lobes of the corolla and inserted on them. The flowers are quite regular. They all have some form of a capsule for fruit, which in most generally split lengthwise. Familiar examples include the garden primrose (not the wild evening primrose), yellow loosestrife, the star flower, pimpernel, shooting star, and the beautiful cyclamens. A few members of the family are slightly luminous in the dark, apparently an attraction to night-flying insects. About 28 genera and over 400 species, mostly from the northern hemisphere, a few in temperate South America and South Africa.
Gentianaceæ—Gentian family. Over 700 species in 70 genera, all bitter herbs, with opposite leaves, quite without teeth and beautiful, sometimes fringed, always regular flowers. In this and related families the stamens are of the same number as the lobes of the corolla, and always alternate with them. Gentian and sea or marsh pinks are our best-known native representatives, while some related plants are medicinal.
There are many other families in this part of the scheme of plant classification that have minor differences among themselves, but agree pretty generally in the number and position of their stamens, their superior ovary, and, on the whole, in the regular flower. Irregular and regular flowers may be recognized at once by cutting them lengthwise through the middle. In regular flowers there would be as much on one side as on the other of the dividing line, and in irregular ones quite obviously more on one side than the other. The character of all the genera in a family having irregular flowers begins to occur here with greater and sometimes exclusive frequency. In the mint family, or Lamiaceæ, nearly all of its 160 genera and over 3,000 species have two-lipped or irregular flowers. The garden salvia well illustrates the type.
The Lamiaceæ or Labiatæ, as they are often called from the two-lipped corolla, are herbs locally, but in the tropics often shrubs or trees. Almost universally they have four-sided stems and opposite leaves without stipules. The flowers may be solitary, much more often they are crowded into various kinds of clusters. The four stamens are borne on the corolla tube, and nearly always there are two long and two short ones. The family is universally fertilized by insects, and some of the flowers are wonderfully adapted for this end. Common examples, besides the salvia, are mint, thyme, skullcap, hyssop, bugle, blue curls, catnip, hedge-nettle, coleus, and Oswego tea. Most of the genera contain heavy odorous oils in their foliage, from which oil of mint, pennyroyal, lavender, rosemary, marjoram, savory and balm are the best known. These volatile oils give to members of the family their characteristic and often very beautiful odors.
There are many other families of plants, some with irregular and others with regular flowers, that appear to group themselves around the Lamiaceæ, all of which agree, in spite of individual differences, in having a superior ovary. The remaining families of the gamopetalæ, however, always have an inferior ovary, usually obvious by the insertion of petals above the ovary, and in the fruit often conspicuous by the remains of the withered calyx still clinging to the top of the fruit. Only two of the scores of families, with inferior ovary and gamopetalous corollas, will be mentioned here, both of which are important.
Rubiaceæ—Madder family. Common examples are the creeping bedstraw, the sweet woodruff, partridge berry, button bush, and bluets or quaker-ladies. All, except one of these, are herbs, but in the tropics the Rubiaceæ are nearly all shrubs or trees. Among those are the coffee, quinine, and ipecac. All the family have opposite leaves (a few verticillate) and stipules, regular flowers, with stamens as many as the corolla lobes, and alternate with them. The fruits are a drupe, berry, or capsule. Over 340 genera and 6,000 species are known almost throughout the earth.
Compositæ or Carduaceæ—The daisy family and the largest and most complex of all the plant families. As the culmination of the scheme of plant classification, they show the greatest perfection in the arrangements for cross-fertilization. For a description of their flower structure, see Chapter I, page 44, and Figures 43-45. Some of the Compositæ have no ray flowers, others are all ray flowers, but the great bulk of them bear both tubular and ray flowers in a single head. This may be single, or more commonly it is arranged in various kinds of clusters. Each head is surrounded at its base by one or more series of usually tightly overlapping bracts, incorrectly called a calyx by the unobservant. The Compositæ include over 900 genera and 11,000 species from all parts of the world. Most of them in America are herbs. Daisy, dandelion, dahlia, chrysanthemum, sunflower, boneset, chicory, lettuce, and scores of other examples could be cited, all herbs. In the tropics the Compositæ are more often trees and shrubs. The family contains many economic plants such as arnica, chamomile, artichokes, inula, and many others.
This all too brief account of the grouping of plants in families, and the sequence of these from the comparatively simple naked-seeded pines, through monocotyledons, the apetalous dicotyledons, followed by the polypetalous dicotyledons, and culminating in the Compositæ among the gamopetalous families, gives us merely a hint of what are the characters upon which plants are divided. While the details are necessarily omitted, the gradual development from naked-seeded plants, wholly at the mercy of the elements, up to those which are marvelously provided with contrivances to insure cross-fertilization, has been traced. Perhaps no other phase of botanical study offers such a rich opportunity as this, for upon the solution of some of the problems of plant classification depends the answer to many questions about the history of the earth and man’s ability to live on it. Certain of these plant families have lived on the earth hundreds of thousands of years before man first came. Others have apparently arisen comparatively recently. Many botanists believe that all the monocotyledons should be placed after the dicotyledons, as the latter may be a more ancient type than the former. How these different plant structures, some very ancient and others more recent, help to show us some of the history of the earth, will be treated, among many other evidences of a plant nature, in the chapter on “The History of the Plant Kingdom.”