CHAPTER II: PLANT BEHAVIOR
1. Light and Its Importance To the Plant
PRACTICALLY all that has been said in the first chapter relates to what plants are, their organs, or what we may call the architecture or plan of their framework. But what they do with this elaborate structure is as important as what we do with a house that may contain every modern improvement but is never a home until these things have been put to use. One of the chief concerns of any architect is to see to it that the house has as much sunshine by day and as attractive illumination by night as possible. Nature, that greatest of all architects, also sees to it that plants get the utmost necessary sunlight, but for a much more important reason than the mere attractiveness of sunshine, be that ever so beautiful. For light, the life-giver of all green things, is so absolutely essential to plants that experiments to grow them in the dark have always failed, and many gardeners now use electric light in greenhouses in order to prolong the short daylight of winter. It is the lack of light that makes celery blanch.
Plants grown in the house inevitably turn toward the windows, even plants growing against a wall turn their leaves away from it—nowhere can one find living green things that do not find the light as surely and persistently as men try to get their food or their mates. Many examples of this could be given and must have been noticed by everyone.
Sometimes seeds germinate under a barn floor for instance, and the puny pale little plantling reaches out slender stems, all of which turn, as a compass turns to the north, to perhaps a crack of light in one corner of the building. We have already seen how the search for light will carry the slender rattan palms of India hundreds of feet to the topmost leaves of the forest. Individual plants, and, as we shall see later, whole forests make desperate efforts to get to the light. We know already, that the struggle for light is just as bitter as the struggle for food by roots. And finally if, as we have many times proved by experiments, plants die when grown in a dark room, what is it that light does for plants and how is a process carried on that everything leads us to think is of the greatest possible importance? Quite obviously it is not the mere beauty of sunshine dancing upon the landscape, as entrancing a picture as that may be any summer afternoon, with the play of sunshine and shadow on the tracery of foliage. That green color of the foliage, the almost universal green of so much of the earth’s vegetation, restful to tired eyes, providing us with the most pleasant shade, has wrapped up within it the secret of just what sunshine does for plants. For under the magic of light acting upon this greenery one of the most important industries in the world, the manufacture of food, is constantly going on.
LEAVES AS FACTORIES FOR THE MAKING OF FOOD
It must be clear enough from the start that to call a leaf a factory for the making of food forces us to decide at once whether this is a mere way of speaking, or whether, incredible as it may seem, anything as thin as a leaf can really produce food. As we eat lettuce, and millions of cattle graze every day, leaves as food producers win handily on that score. But to understand how food is produced in such a tiny factory demands that we walk about in it for a bit, study the inside of it and especially its many small chambers within which is not only the machinery, but some of the finished product stored up for later use.
Unlike modern factories there are many entrances, from any one of which we can begin our tour of inspection. On the under side of nearly all leaves and on the upper side of some there are scores or even hundreds of small pores called stoma, so small that only with a microscope can they be seen. These entrances through the factory wall, are carefully guarded by a pair of watchmen whose business it is to see neither too much dry air gets in nor too much of the product of the factory gets out. They see to it, also, that waste products are thrown out at the proper time. These watchmen, or guard cells, as they are called, are constantly on the job, work almost automatically, but their chief function is connected with the proper ventilation of the place, and will be discussed later under “How Plants Breathe.”
Once past the entrance it is obvious that we are in one of the strangest of all factories, for none of the rooms are truly square or oblong and their irregularity as to outline would drive your average foreman into profanity. Yet they are certainly divided into distinct classes, at least as to size and as to what the rooms contain. Some are apparently filled with nothing but air and have direct connection through the stoma with the outdoors. These are called intercellular spaces. Others, and these are most important, are filled mostly with the green coloring matter that gives the leaf its color. This substance is known as chlorophyll, its individual units as chloroplasts, or literally, chlorophyll bodies. Quite independently of these chlorophyll cells or rooms, or the intercellular spaces which correspond to halls, there are some large and many small tubes. These are the veins of the leaf and their finer branches and by their direct connection through the stem to the roots, serve as the ducts through which some of the raw materials are brought into the factory.
This green coloring matter or chlorophyll is perhaps the most important substance in nature. Without it all except a very few plants would die, and even in those beautifully colored leaves like coleus or caladium chlorophyll is always found, but in these colored leaves it is merely obscured by other coloring substances. It is in the chlorophyll that the ability resides to take the inorganic substances through the roots or from the air, and by the aid of sunlight transform them into organic substances like starch and sugar. Nothing else in all nature can do it; without this faculty, which the commonest green leaf possesses, the earth would prove uninhabitable within a single year. Just what chlorophyll is chemically is not yet thoroughly known, but the thing of chief interest is that it is hardly ever found in parts of the leaf not exposed directly to the sunlight, and that during the autumnal coloring and before the fall of the leaf chlorophyll is carried to other parts of the plant, and quite possibly stored for use the following season.
While the composition of chlorophyll is not surely known, iron is certainly one of its constituents, as plants deprived of iron lose their green color. It also is known to contain oxygen, carbon, hydrogen, and nitrogen, but merely to catalog what we know about its make-up does not tell us that it is a living green substance and that sunshine sets it in motion. Just exactly how light acts on chlorophyll no one really knows; we merely know that it does so act and that the result is one of the marvelous secret processes of nature, perhaps like the secret of life itself forever hidden from man. In our tiny factory, then, we have raw products coming from the roots and through the stoma from the air; machinery of the most efficient type, for chlorophyll works night and day, and constantly renews itself while producing the finished products; energy from the sun; and finally the complete manufactured products which are foods in the shape of starch and sugar. During the growing season there is no banking of the fires, no stoppage of this most important of all industries, no strikes or lockouts. Each part of the whole works smoothly and with the nicest precision—in fact so perfectly does this process keep on going, so complete is the orderliness of the place, and so regular are the completed products turned out, that no modern factory manager or workman but can learn something from a rather close study of this smallest but most efficient factory in the world.
Some of the raw products are delivered to the leaf from the roots where they have been absorbed by another process that will be considered a little later. These consist of water and the inorganic substances dissolved in it, popularly called sap. Carbon and oxygen come mostly from the air, sometimes separately, more often in the form of a combination called carbon dioxide which is one of the chief constituents of the gas thrown off by man as he breathes out. Now these inorganic substances, contained in the sap or derived from the air, are literally mixed by the chlorophyll and form, always with the aid of sunlight, substances known as carbohydrates, the commonest example of which is sugar. Some form of sugar is one of the earliest results of this process, but sugar is quite easily dissolved in the sap which has contributed to its manufacture, and the excess sugar is thus removed. Otherwise it would clog the machinery and prevent the production of fresh supplies. This first step in the manufacturing process has not inaptly been called photosynthesis, the meaning of which photos, light, and synthesis, combining by means of, suggests in a word the necessity of light and the combination of the inorganic substances mentioned above. Of course this process of photosynthesis is not as simple as the brief account of it suggests, for it is actually a complicated chemical process only part of which is yet understood. It is fairly certain that it goes step by step; it is quite certain that the beginning is inorganic and the end organic compounds like sugar. Something is known also of the wear and tear on the chlorophyll, its waste products, and how it keeps itself not only fit but provides for its own constant renewal. One of the excess or by-products in this initial manufacture of sugar is oxygen. This is either used in other ways by the plant, or more generally it is thrown off through the stoma into the outer air. Oxygen, as one of the necessary constituents of the air that man breathes in, is thus thrown off, while, as we have seen, carbon dioxide, a poisonous gas which we breathe out, is a necessity for this manufacturing process in all green plants. Hardly any trick of nature so completely fulfills the wants of animal and plant life as this mutual exchange of by-products—in the case of animals it is the waste of respiration, in plants it is the wastage of sugar making and some other changes that go on in the plant just after this stage.
The amount of sugar made, carbon dioxide taken in, and oxygen given off by this process suggests that while leaves may be very tiny factories they are among the most efficient in the world. Assuming an area of leaf surface equal to about a square yard the amount of sugar made would be about one-third of an ounce in a day or nearly three pounds in a single growing season. Carbon dioxide withdrawn from the air would average from the same area of leaf surface about two gallons a day or over three hundred gallons for the season. As an equal amount of oxygen is given off by the leaf, it becomes clear that as all of this interchange must go through the stoma the functioning of these and their guardians must be nearly one hundred per cent perfect. As we shall see a little later, they perform still other duties with even greater perfection. When we stop to reflect what an absurdly minute fraction one square yard of leaf surface is to the total leaf surface in the world, we come to some realization of the gigantic proportions of this process of manufacturing sugar and exchange of gases mutually useful to animals and plants. While in the United States most of the leaves fall in the autumn, the great bulk of the vegetation of the world holds the greater part of its leaves all the year, notably in the vast evergreen forests in the north, and of course practically all tropical vegetation. Chlorophyll in such places works continually and what the total of sugar production may be no man can even guess.
Sugar, although the first step in the process, is not the final one, and the leaf has still other tasks to complete. Some of the sugar is used up in the process of renewing the chlorophyll, some of it is moved to other parts of the plant where in sugar cane it forms the world’s chief sugar supply; but the remainder is transformed into starch, a substance that is not dissolved by the water of the sap, and is therefore capable of permanent storage either in the leaf itself or in other parts of the plant, notably in the tubers of the potato, the solid part of which is nearly all starch. The conversion of sugar to starch, which is really a means of contriving to properly store the product of the factory, is done by certain ferments known as enzymes. Just what enzymes are or even how they work is not well known, but apparently they have the faculty of converting certain substances like sugar, and in the process they neither use up nor materially change their own composition. It is certain that the conversion of sugar to starch is an elaborate chemical process, but it is accomplished by these enzymes, the very existence of which has only recently been discovered. Enzymes not only do this, but they convert starch which is insoluble into a kind that may be dissolved and thus carried to different parts of the plant. Upon this power depends the storage of starch in roots, tubers, seeds, or wherever else it is found in the plant, and it is of course upon this power man depends for the food supply of the world. Wheat or corn, potatoes, rice, all the foods that are rich in starch produce none in that part of the plant harvested by man. All of it has come by the process which is only sketched in its briefest outlines in the foregoing paragraphs. All of it must come from that green coloring matter of nearly all plants which, while mostly confined to leaves, is not always so. And wherever chlorophyll is found this process goes on even in the simplest plants. Because it is so overwhelmingly a characteristic of leaves and, as we have seen, leaves are the one organ of the plant upon which man pins his only hope of future food supply, the leaves of all plants may be truly likened to a factory the work of which is never ending, the product of which the leaf will never use, but the result of which has far-reaching consequences to us all.
EFFECT OF LIGHT AND DARKNESS ON INDIVIDUAL PLANTS AND VEGETATION AS A WHOLE
Now that we understand the importance of light to all except a very few plants, and its very close relationship to the green coloring matter of all leaves, many things about the arrangement and position of leaves, and indeed of the whole plant, may be understood, which, without this knowledge, seems the result of mere caprice or chance. It would seem as though the habit of plants growing toward the light, and against the pull of gravity, a character almost universal, no matter from what mountain declivity or rocky cliff it may spring, might be the result of the “pull” exerted by light on the green coloring matter in the leaves. While light does aid in plants having a generally erect habit it is not the cause of it, as we have many times proved by experiments. As a seed sprouts and the roots go down into the earth, the shoot, before it has broken through the surface and while still in the dark, always grows upward. This property of growing in two opposite directions at the same time, the roots always with gravity and the shoot nearly always against it, is known as geotropism. In the case of vines or other trailing plants there is the same tendency exhibited, even though the plant is not erect. We must think of geotropism as a growth habit of all plants, not caused by light, for it has been shown to act in the dark, but of the greatest advantage to all plants in their initial start toward the light. If this were not the case, it may be imagined into what chaos the vegetable world would be thrown. We are so accustomed to roots going down and shoots going up that we are not apt to think of it as the result of two antagonistic growth habits, the true cause of which is not understood, the result of which is common knowledge. Geotropism is one of those mysteries with which the book of nature is crowded, and merely to describe it and realize its force is by no means to arrive at its true inwardness.
But, quite independently of this peculiar growth habit, the stems and often whole plants do show response to light and many times the response, in its effects, cannot be distinguished from geotropism. Perhaps the most homely illustration of this is the common house geranium which, no matter how often it is turned, always grows toward the window, and if not turned at all becomes hopelessly lopsided, with the leaves all bending sharply toward the light. Trees growing on a cliffside, while always growing upward, nearly always may be seen bending away from the cliff where light is scarce and toward the unobstructed light. The position of hundreds of twigs and branches on any tree have been dictated by their exposure to light, and the habit of practically all trees in the forest of being clear of branches for many feet from the ground is another illustration of the profound effect of light. In the latter case the taller the trees the farther from the ground are the first branches, and in the big trees of California the first branches are frequently over a hundred feet from the ground. In their young stages all these trees were furnished with branches, the leaves of which in their day performed their appointed tasks. But in the strife and hurry of the crowns of the forest to overreach their neighbors these lower branches, from the bottom upward, gradually die off. So inexorable is the plant’s demand for light, that these lower branches, in spite of being nearest the source of their food from the roots, are doomed to be killed. Nature plays no favorites and these lower branches, once the pride and support of the young tree, are ruthlessly dropped off when they can no longer play the game. This wholesale slaughter of lower branches in a forest, more complete than any pruning by man could ever be, gives us, if the story of the factory leaf has not already done so, some conception of the part played by light in the plant world.
The shade of certain trees is so much denser than others that they have been planted for this purpose, notably the horse-chestnut and Norway maple. Foresters have long recognized this difference in trees and it would be strange if nature had not taken advantage of it also. If certain trees can still maintain themselves in the forest without producing a dense crowd of leaves, such as the silver maple for instance, they would have a decided advantage over a tree like the sugar maple which casts a much denser shade. A walk through any forest will show scores of examples of trees that live and produce seeds by virtue of the fact, not that they demand all available light, as their more vigorous neighbors do, but that by a compromise, by an almost diabolical cunning, their light demands, and of course their leaf exposure, have been cut down to a point where the tree can grow in a place impossible for trees that lack this ability. It is, of course, not a trick which any individual tree can perform at will. Rather is it a characteristic found in all individuals of certain kinds, where the comparative disadvantage of making less food and having less leaf exposure is more than overcome by the enormous advantage of being able to fight their way into a forest that would otherwise be impossible for them. We shall see, in the chapter on Plant Distribution, how this peculiar response to light has had effects of considerable significance upon forests, particularly after forest fires, lumbering, or other disturbance of the natural conditions. Trees in the forest, and the shrubs and herbs under them are not the quiet stately things about which the poets are so fond of singing. They are places, on the contrary, of intense warfare, and perhaps some of the greatest casualties occur in the battle for light.
Leaves, as being the most directly involved in the matter of utmost exposure to light, show the greatest amount of response to it, by their shape sometimes, by their position nearly always, and very often by the character of their leafstalks. In many herbs the first young leaves are relatively short-stalked, while as the plant grows upward the lower leaves are progressively longer stalked, which is a direct response to the fact that the upper leaves take their full share of light, leaving little or nothing for the lower ones. To avoid complete shading their leafstalks are often many times the length of their more fortunately placed neighbors above them. In those plants like the garden primrose or common weedy plantain, which bear all their leaves in a close cluster or rosette at the level of the ground, we see an almost fiendish cleverness in their earlier and later habits of growth. When the leaves first start, as they nearly always do among grasslike vegetation in which these plants usually have to fight for a chance of life, the leaves grow straight up, so that they may get above the level of the surrounding grass. Once there, and the precious light an assured fact, they gradually flatten out their leaves to form a rosette, of course cutting off the light from the grass about them and killing it just as certainly as though it were pulled up by the roots. Hundreds of different kinds of plants do this, apparently with the utmost cruelty to their inoffensive neighbors, with whom they start upon nearly equal terms in the race for life. If they began at once to spread out their rosette while it was still in its small spring state, the upward pointing grasses would smother it, and as if in anticipation of this the leaves grow up with the grass, only to flatten out when the proper time comes for them to show their true colors.
Light not only affects leaves in their habits of growth but it actually causes movements in some leaves which are as regular as clockwork. The best known cases are those in the pea family and wood sorrels, all of which bear compound leaves. During the day these leaflets are spread out in the ordinary way and catch the light, but at sundown, as though this were a quite useless exertion for the night, they fold up and the leaf “goes to sleep.” On cloudy days they partly fold up, as if in recognition of the fact that for their business of getting light it is an off day; but also if the sun comes out they hurriedly expand their leaflets. It is not yet certain whether these apparently intelligent movements of leaves in relation to light are of any real advantage to the plant as a whole or not. They are surely one of the most interesting things to watch and may be seen in locust trees and wood sorrel any night.
Just as we can have too much of a good thing, it is possible for plants to have too much direct sunlight. In open spaces, where the struggle for life centers not about the fight for light but over other matters, we find leaves actually protecting themselves against too much exposure, and by a variety of ingenious ways. The texture of the upper or lower side, the kind of hair growing on their surface, and the number and size of their pores, are the most usual ways of leaves arming themselves against an oversupply of the one thing that their neighbors in the cool forest fight to the death to obtain. There seems to be a fatality against which plants, like ourselves, are nearly helpless. Their attempts to overcome it, again like our own struggles against an apparently overmastering fate, develop those characteristics that insure survival to the fittest, death to the puny or unaccommodating.
We could hardly leave the subject of light and plants’ relation to it without mentioning, perhaps, the most remarkable case of adaptation to peculiar light conditions. All those aquatic plants that grow beneath the surface of the water need and get much less light than ordinary land plants. But from the island of Madagascar comes the lace leaf or water yam, which grows in quiet pools that are mostly in the depths of the tropical forest. Add to the dense shade cast upon the gloomy surface of such ponds the amount of light naturally lost in its passage through the water, and we get some notion of the singularly secluded home of this aquatic plant. What, now, is nature’s response to these peculiar conditions? How do the leaves of this well-shaded inhabitant of quiet pools behave? Their leaves are about a foot long and three or four inches wide, quite unnecessarily large for a submersed aquatic, but they consist wholly of veins. There is no “meat” to the leaf, none of that soft, green tissue so familiar in ordinary leaves. The conditions under which it is doomed to live almost seem as if it recognized the futility of having a broad expanse of the usually constituted leaf blade to expose to a light which is not there. It is significant that this skeletonized condition is permanent, the leaf functions much as ordinary aquatic leaves do, but its network of quite naked veins almost seems a mute protest against its fate. The delicate, lacelike “foliage” of this aquatic adds a touch of beauty to one of the most curious plants in the world.
2. How Plants Get Their Food and Water From the Earth
If we could stretch an apparently impervious membrane, like the inner white skin just inside an eggshell, or a piece of parchment, and so form a wall through the middle of a glass box, and then pour into one of the compartments pure water and in the other a mixture of water and molasses, a very curious result would follow within a comparatively short period. We should find that presently there would be a gentle filtering of the water through the membrane toward the molasses water, and similar gentle current in the other direction. In other words, fluids of different density, if separated by a membrane, tend to equalize each other. This equalization may not be very rapid, and at first it will be more speedy from the less dense to the more dense, but eventually it will make the different fluids of a common density. This purely mechanical property of the equalization of fluids separated by a membrane is known as osmosis, and it is upon the possession of the equipment necessary for this that roots depend for getting food and water from the soil.
In our discussion of roots in Chapter I, we found that they end in very fine subdivisions, which are themselves split up into practically invisible root hairs. These root hairs are the only way that roots can absorb the food and water in the soil, and they are able to do this because they are provided with a membrane which permits osmosis to act between the solution inside the root hair and the water in the soil. The solution in the root hair is mostly a sugary liquid, some of that surplus sugar made in the leaves, and it is denser than the soil water, so there is apparently nothing to prevent an equalization of the liquids on different sides of the membrane. If this actually happened, as it would in the case of the simple experiment noted above, then roots would exchange a fairly rich sugary liquid for a much more watery one, and we should find that plants did not get their food from the soil, but really have it drained away from them by osmosis. But nature has a cunning device for stopping such robbery, which is prevented by the membranes of root hairs being only permeable to the extent of letting water in, not permeable enough to allow sugar to escape. As we have seen, osmosis is a purely mechanical process which, if left to operate without interference, would not aid but injure the plant. Surely, nothing with which plants are provided is so important to them as this delicate membrane of the root hairs which, while allowing osmosis to act in a one-sided fashion, preserves to the plant the sugary liquid that alone makes the absorption of soil water possible.
As root hairs are very much alive and work constantly, they must be provided with air, without which no living thing can exist. And here, again, it seems as though nature, with almost uncanny foresight, had deliberately planned for this requirement of roots. And, in this case, not by interfering with a physical process by an adjustment of plant structure, but by the arrangement of soil particles and the way in which water is found in all soils. Soil particles, even in the most compact clay, do not fill all the space occupied by the soil as a whole. There are tiny air spaces all through the soil, which insures a constant supply of fresh air. That is one reason why gardens are cultivated, to see to it that plenty of fresh air is allowed to permeate the soil. Around the finest soil particles there is always an almost incredibly thin film of water, which is renewed as soon as it is lost by its absorption by the root hairs or by evaporation. This renewal of the water film is itself a mechanical process, called capillarity, best illustrated by putting a few drops of water on a plate and placing on them a lump of sugar. The water will spread all through the lump of sugar in a few seconds and the capillarity that forces it up through the lump is the same as sees to it that the tiny film of water surrounding the finest soil particles is constantly renewed from the lower levels of the soil.
Little do we dream, as we walk over the commonest weed, that buried at its roots are these delicate arrangements for securing food and water. Osmosis allowed to act so that the “exchange” of liquids is all to the advantage of the plant, capillarity providing a constant water supply, and the very piling together of the soil so contrived that the life-giving air filters all through it—does it not seem as if all this were, if not a deliberate plan, certainly a more perfect one than mere man could have devised?
If you will turn back for a moment to the beginning of the description of how plants get their food, you will find that in osmosis the weaker liquid tends to permeate the denser one more rapidly than the denser one does the weaker. As we have just seen, the sugary liquid in the root hairs is denser than the soil water outside, and, furthermore, none of it is allowed to escape. This comparatively greedy process of taking everything and giving nothing results in a constant flow of soil water into the root hairs. When the flow of liquids in osmosis is not at once equalized, a gentle pressure is brought to bear to make them so. This is what is called osmotic pressure, and it is this pressure that forces the absorbed liquid through the roots and part way up the trunk of even the tallest trees. While we have just said it is a gentle pressure, that is true only in the case where the osmosis has free play, and the pressure is stopped with the perfect mixing of the two liquids. But what if they can never mix? What may not the accumulated osmotic pressure amount to in such a one-sided process as goes on in root hairs with everything coming in and nothing going out. Cut-off stems, with a pressure gauge attached to them, indicate that in some plants the pressure is from 60 up to 170 pounds!
Another result of this pressure is that it keeps leaves and the fleshy stems of plants in their ordinary position. The actual solid part of nearly all leaves is scarcely 5 per cent of their bulk and all the rest is water. The constant pressure of this water from the roots is sufficient to keep leaves comparatively stiff and rigid, how stiff is quickly realized if the pressure stops and the leaf wilts or withers. Sometimes this osmotic pressure, particularly during rainy weather, becomes so great as to cause injury to the plant, the splitting of tomatoes and occasionally of plums, being due to it. This osmotic pressure, together with the extra pull given by the leaves, is sufficient to account for the rise of water to the tops of the tallest trees. The tallest trees in the world are certain kinds of blue gum in Australia which frequently reach a height exceeding 300 feet. What the combined osmotic pressure and leaf pull must be to carry such a heavy thing as water to such a great height is easier to imagine than to calculate.
The root hairs, then, by the process already described, absorb the water from the soil, but plants can no more live on water alone than we can. As we have seen, the membrane in the root hairs cannot allow the passage of even the tiniest particle of solid matter. In fact the root hair itself is so small that it can only be seen through the microscope, and of course the membrane is smaller still. Plant foods, then, can never be solids, but must always be such materials as can be dissolved in water. The chief of these are chemical substances, such as lime, potash, nitrogen, magnesium, phosphorus, sulphur, and iron. Hydrogen is also necessary, but as this makes up half the composition of water, there is a permanent supply of that provided by the soil water. These things make up the great part of plant foods taken in through the roots, and it is from these that leaves, by a process you already understand in its essential details, manufacture sugar and starch.
But neither starch nor sugar, important as they are to the plant, and absolutely necessary as they are to us, are the only things made by plants. Leaves may well be called factories, but plants are themselves the most wonderful chemical laboratories, beside which any built by man are as play-things. For plants, by processes too complicated to be explained here, work over their accumulation of starch and sugar, recombine some of their constituents, and store up in various parts of the plant the results, which are often such food ingredients as protein. This is the really essential food substance in wheat, as it is in eggs and meat. No chemist has ever succeeded in making a single scrap of it, yet it is such an everyday occurrence in practically all plants that it, with starch and sugar, forms the great food supply of the world. Not protein alone, but all the amazing plant products like the oils from the olive and the resin from pine, rubber, the drugs of plant origin, even tobacco—all these and hundreds of others are made by plants from those few simple foods absorbed through the roots, literally pumped up to the leaves and there, under the magic of sunlight, combined and recombined, worked over and changed utterly in their make-up. Nothing could be more perfect than the marshaling of forces and contrivances to secure the result; let there be even the least bungling, and for us the world would cease to be worth fighting for.
Nor does the work of plants stop here. If it did, they would be not unlike a commission merchant who had gathered from the four corners of the earth a supply of eggs only to find he could not or more likely would not sell them all at once, and yet had failed to provide himself with proper storage. Plants, too, have times in their life when adequate storage is necessary for them. So true is this that unless there is food enough stored in seeds to give a start to seedlings before their own roots begin to work, they would die almost at once. In seeds and in many nearly dormant parts of plants these foods are stored away for future use. The tubers of potatoes and all our root crops, like beet and parsnip, are common examples of this. Even the manufacture of wood in the trunks of trees is a storage appliance on the part of the plant, for wood is just as much one of the food products of a plant as wheat or rice.
3. Borrowing From the Living and Robbing From the Dead
With such a beautifully perfected mechanism for getting food it might seem as though all plants would be satisfied to lead that life of independence for which they are so splendidly equipped. Some of them, however, are like men in one respect: there seems to be no end to the chase after getting something for nothing. Those that stand on their own roots, get their food honestly, and take nothing for which they do not make prodigal returns, make up the great bulk of the vegetation of the earth. Their independence has dubbed them with the title

Indian Pipe. (Monotropa uniflora). A saprophytic plant inhabiting rich woods in eastern North America. (Courtesy of Brooklyn Botanic Garden.)

The Partridge Berry (Mitchella repens), a trailing vine of northern forests. (Courtesy of Brooklyn Botanic Garden.)

Rafflesia. One of the Strangest Products of the Rain Forest. It consists only of a giant flower, the largest in the world, which is attached directly to the roots or stems of relatives of the grape, upon which it is parasitic. (After Kerner and Oliver. Courtesy of Brooklyn Botanic Garden.)
autophytes, literally solitary or self-providing plants, and this thrifty mode of life is called autophytic. But a few kinds of plants, actually many millions of individuals, have more devious ways of getting their food and provide strong contrast to their sturdier associates.
These baser modes of life appear to have been rather insidiously developed, as though there had been some hesitation at even the smallest departure from the normal. Of course we must not forget that plants, while living things, are never reasoning ones, and that good and evil and all other qualities that are ascribed to plants are perfectly foreign to them. Throughout this book, and in many others, the habits of plants are spoken of as base, for instance, or good. What is actually the fact is that nature works in truly marvelous ways, and to our reasoning faculties these adjustments seem clothed with attributes they do not really possess. But the description of them in the terms of our everyday speech, the translation of their behavior into the current conceptions of mankind, does so fix them in our minds that they cease to be “just plants,” and we no longer put their habits in the category of those interesting things that nearly everyone forgets.
One of the first signs of departure from the usual methods of getting food is the association of certain minute organisms at the roots upon which plants, otherwise autophytic, depend for aid in securing nourishment. This characteristic is fairly common, notably in all the plants of the pea family, such as peas, beans, locust trees, vetch, clover, and hundreds of others. If the roots of any of these be examined, it will be seen that attached to the smaller divisions of them are small tubercles from the size of a pinhead to a pea, depending on the kind. These tubercles or galls are caused by and infested with bacteria, the smallest of all plants. The bacteria have the extraordinary power of changing nitrogen into nitrates, which is the only form in which nitrogen can be absorbed by roots. Not only do they accomplish this, but excess nitrogen is stored in the roots by the same agency. It is this fact that has resulted in the planting of vetch and kindred plants for soil enrichment, as each year there is a residue of nitrogen left in their roots and by so much they add plant food to the soil. For hundreds of years farmers have done this, but only quite recently have we known why they did so. The occurrence of bacteria or microbes at the roots of plants is much more common than was formerly supposed to be the case, and many other plants than those of the pea family depend, at least in part, upon them in getting food from the soil. While not wholly autophytic, such plants do make some return for what they gain, as some of them at least pay dividends in extra nitrogen, and all of them provide opportunity for the bacteria to live. The latter play an important part in populating the soil, which is not the comparatively sterile thing it appears to be. Actually it is infested with organisms that play a mighty, if rather inconspicuous, part in the work of preparing the soil for plant growth. These organisms are so minute and the chemical nature of their work is so complicated that merely to mention their existence must suffice here. This close association of certain roots and bacteria, which, as we have seen, is of mutual advantage, is known as symbiosis. It is really only a kind of exchange, not unlike the storybook community that helped out by taking in each other’s washing. Unlike that community the association between the two works to the actual advantage of both, but the process is undeniably a step away from those wholly autophytic plants which live free and independent of such aid.
A much more gruesome habit of certain plants is their reliance for food only upon the dead. In the Indian pipe, some kinds of shinleaf, and in many other plants their roots and root hairs are changed or often nearly lacking, and we find them growing only on the dead bodies of other plants. One peculiarly repulsive characteristic of such plants is that they secrete at their roots a substance that hastens the decay of the dead, and, as if this were not rapid enough, there are associated with them certain kinds of minute fungus organisms that also speed up decomposition. Plants with this charming mode of life are known as saprophytes, literally sapros, rotten, and phytes, plants. “Rotten plants” they may be in their mode of life, but the pearly white stems and flowers of the Indian pipe have a certain ghostly charm, an almost statuesque beauty among the normal greenery of the gloomy dark woods in which they always grow. It is not without significance that Indian pipe bears no leaves, has none or almost none of the life-giving green coloring matter which we have seen to be the almost priceless possession of plants which lead a different, and perhaps a better life. The great bulk of saprophytes bear no leaves, and some only partially wedded to the habit appear to be midway between bearing normal green leaves and bearing none, or much reduced ones that are quite unlike the busy factories we know normal green leaves to be. Plants with this method of getting their food, must of course grow in places where dead and decaying vegetation is plentiful, and often as such soil is turned up there may be noted a peculiar dank odor, suggestive not only of its origin, but of the fact that these “rotten plants” make their home in it. Some of our most beautiful orchids grow in this fashion, but even there, in spite of flowers that for beauty of form are without rivals, the plants have no green coloring matter in their leaves, which are often reduced or even wanting altogether.
It might almost seem as if demoralization, so far as food habits are concerned, had reached its lowest point in these plants that literally rob the dead, but there are still lower depths to which certain plants have been reduced. This consists of robbing the living, and such plants are called parasites, a word perfectly familiar in other connections. Parasitic plants have no roots, but attach themselves to the roots of other plants, somewhat generously called hosts, from which they derive their food. The best known case is the common Christmas mistletoe, and the dodder (Figure 68), but there are hundreds of others. Nothing in all the realm of plant life so perfectly fits the action to the word as plants of this type, flourishing when the host flourishes, dying when it dies. Producing flowers and seeds, and often, by an irony of fate, perfectly green leaves, they are nevertheless the most debased of all plants in their mode of life.
These successive steps in the degradation of food habits, are not always the clean-cut things they might be inferred to be from the foregoing. There are many intermediate stages; it may even prove to be the case that some plants are wholly autophytic at certain stages of their life, and slip partially into more devious practices at other stages. The whole affair is not yet thoroughly understood and may well be the result of competition, as it is quite conceivable that if the getting of food in normal ways became difficult or impossible plants may have had to resort to other methods.
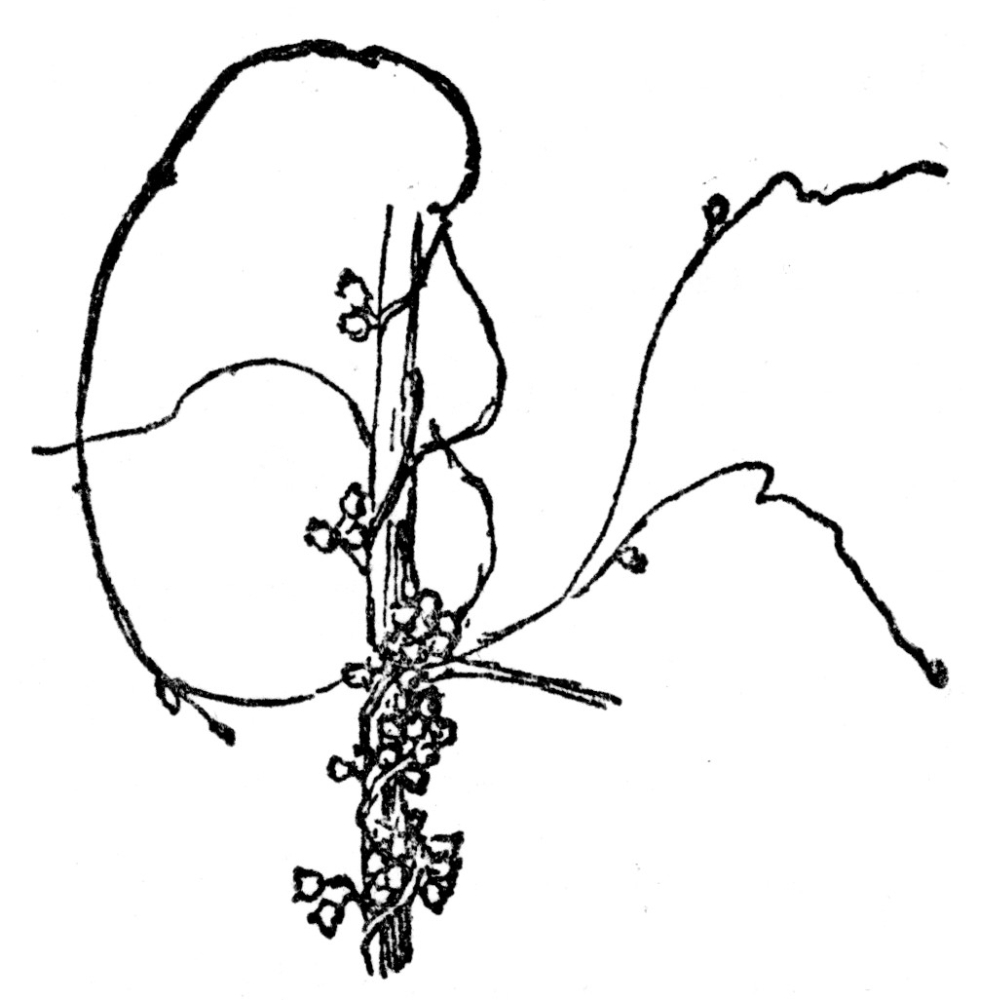 FIG. 68.—THE DODDER A leafless parasitic vine which steals its food from the plants to which it is attached.
FIG. 68.—THE DODDER A leafless parasitic vine which steals its food from the plants to which it is attached.
4. What Plants Do With Water and How They Breathe
Some one has said that one day without water would make men liars, in two days they become thieves, and after the third or fourth day they would kill to get water. In the Army Records at Washington is a report of one of our expeditions, which in chasing Indians got lost in a desert, and in which the soldiers fought among themselves for even the most repulsive liquids. It hardly needs these gruesome examples, however, to confirm what everyone who has ever been mildly thirsty knows, that water is an essential for all animals, and that to be without it is to suffer torture. Air of the proper kind is just as important, and because its absence or impurity causes more sudden agony and a quicker death, the need of it is that much more acute. Plants rely even more upon these two essentials of life, and in getting them they behave in ways just as ruthless as do men who are suddenly deprived of either of them.
As we have already seen in “How Plants Get Their Food and Water from the Soil,” the water is the carrier of the food elements from the soil, but water as such does much more for the plant than act as a carrier. Osmotic pressure, a never-ending pump, keeps sending up a steady stream of water to the limits of its power. In everything except trees it seems fairly certain that this pressure is sufficient to drive water into the remotest leaves. It finally reaches these tiny rooms in the leaf about which we read in the account of Leaves as Factories. And just here a very curious thing happens. Each room is, as we have seen, a very busy place, crowded with all the necessary equipment to make sugar, and yet there is still room for water which is just as necessary as the other fittings; in fact so necessary is it that the whole interior of the room is bathed in water. This irrigation system works so well that the walls of the room literally bulge with the pressure of the water in them. If they did not—a condition known as turgor—the plant would at once wilt, and if no new supply came it would wither and die.
But water cannot stay in this condition of pressure and stagnation for even a brief period. That would be as if a leaf were like a toy balloon which, after inflation, had the entrance pinched and so remained inflated. And while we have all along spoken of factories for making sugar, and pressure pumps for forcing up food and water, it must never be forgotten that this marvelously adjusted mechanism is a living thing. Constantly growing, even producing their own means of falling in the autumn, leaves must be thought of as living machines, just as we are still more highly developed machines. In other words the accumulated water in the cells of the leaf must be removed, after it has served its use, and replaced by fresh supplies. The removal is carried on by its evaporation into the halls, or, in the more precise terms of our account of leaves as factories, into the intercellular spaces. It will be recalled that these are connected with the outside air through the pores or stoma. When the air outside is hot and dry it might easily suck out by evaporation all the water vapor in these intercellular spaces and wilting follow at once. This would actually happen if the guard cells, already mentioned, were not constantly on the job. They control the size of the opening just as certainly as a steam valve does, and maintain, with a few exceptions, just the proper amount of water loss not only to maintain turgor, but to see to it that transpiration, as this process is called, goes on rapidly enough to insure fresh supplies of water being sent to the leaf. The opening and closing of the stoma by the guard cells is a nicely balanced operation dependent upon root pressure, turgor, and atmospheric conditions. Guard cells have, because of this, been much studied in spite of the fact of their microscopic size. We now know that they allow greater openings during the night and reduce them during the day. When we reflect that the constant removal of water in the leaf, both as such, and as the only carrier of food supplies from the roots, depends in such large measure upon the functioning of these guard cells, then we come to some realization of their importance to the plant.
They do not always work unaided, for in many places the transpiration, even with their best efforts, would exceed the rise of water in the plant and death must follow if such a condition exists for long. This may be the case in certain bog plants, where, even with their roots in the water, they actually are in danger of drying out because the composition of bog water makes it partially unfit for most plants. And, again, in very open dry or windy places, such as deserts or the mountain tops above timber line, the actual supply of water may be insufficient. Many plants growing in such places have their leaves, particularly the under surfaces of them, clothed with various kinds of hairs. These may be quite velvety or cottony, but in any event, either by their texture or their color, they tend to reduce transpiration. An extreme case is a desert plant from Arizona where the whole leaf surface is covered with an ashy gray velvety coating, which, of course, absorbs less heat than a normal green leaf, and in addition there are much fewer pores through which transpiration could be carried on. In ever so many leaves nature has provided them with a thick coating of hairs in early spring, which they lose later in the summer. Shrubs and herbs, especially those that start earlier than the trees under which they grow, very often may be found with a dense woolly or silky covering in early spring. As the shade becomes denser and the need of the protection less, the wool or silk is shed, sometimes completely. Some of the most conspicuous cases of this are certain kinds of our common shadbush, which in April are covered with a beautiful grayish-white silky coat, but by August are practically the ordinary green color of other leaves. The hairy covering of leaves is well worth observation, as it may hide not a few facts about transpiration and, in some leaves, has had much to do with their preservation from grazing animals. Some, like the common mullein, are never touched, and may be found standing like sentinels in fields otherwise cropped short.
In many leaves there is conflict between those forces that result in the leaf getting the utmost possible exposure to light and those that prevent too rapid transpiration. On the one hand there is the absolute necessity for light, on the other the ever-present danger that the response of leaves to this necessity will result in a transpiration rate too rapid to be held in check by the guard cells. The compromise between these two forces, each pulling in opposite directions, gives to some leaves a series of movements that are among the most interesting things in nature. One of the most marked examples is the common wild lettuce, a weedy plant of our roadsides introduced from Europe. In bright sunlight the leaves are turned so that the edge of the blade faces upward, and the surface is thus protected from the direct rays of the sun, but during cloudy weather or in the shade the leaves turn into the ordinary position of most foliage leaves. It is difficult to avoid the inference that photosynthesis, which, as we have seen is an absolute necessity to the leaf, is in the wild lettuce retarded by transpiration, to avoid the too rapid rate of which the leaf is turned on edge. In this plant the leaf base, as though to be ready for whatever change transpiration or photosynthesis may demand, is so attached to the stem that such changes are made with the least possible delay or wrenching. In one of the many kinds of blue gum trees of Australia all the leaves turn one way in the light, and another in shade or on cloudy days. Ever so many plants have partial movements of their leaves, a good many of which are in response to these opposing demands, one pulling the leaf into the greatest possible light, the other holding it away from that condition. There are other movements of leaves, of parts of the flower, or even of the whole plant that are not so certainly the result of the conflict between light requirements and the necessity of conserving water supply. They will be considered presently.
While most plants are well provided with methods of losing water, so well provided in fact that in very hot or very long dry periods it is a common sight to see many plants literally panting for more water, there are some apparently more cautious individuals, who reverse this process. All throughout tropical America hundreds of relatives of the pineapple have their leaves so formed and arranged that they catch and hold considerable quantities of water. In one kind, called Hohenbergia, the long leaves are joined together toward their base into a water-tight funnel, which will hold a quart or two of water over a period of drought. In Africa the extraordinary traveler’s-tree, a giant herb growing twenty to thirty feet tall, has the overlapping leaf bases so arranged that they hold many gallons of water. And we have already seen how the giant cactus of our own Southwest will hold 125 gallons. The most remarkable case is the Ibervillea from the deserts of Arizona. In riding over this country one may find objects that look not unlike a burned pudding, about two feet in diameter and nearly as high. From the center comes a delicate stalk with the finest feathery foliage and tiny flowers. Of roots there appear to be almost none, and these curious objects, which are very hard and woody, might almost be taken for stones. But they are actually plants not distantly related to squash and pumpkin, and one of them collected years ago and brought into a museum behaved in quite the most thrifty fashion of any plant yet discovered. It was carefully cleaned and put in a museum case and locked up as a curiosity for the wondering public to gaze at. But suddenly, almost miraculously, it sent out its delicate growth which grew its appointed time and then withered. Imagine the astonishment of the curators of this museum to find it doing the same thing the next year, and the next. Finally after putting forth its shoot for five years it actually died and is now a peaceful museum specimen. No other such case of water storage is known, but thousands of plants have this remarkable ability to a less degree, all in response to conditions that would mean destruction to plants not so providently equipped.
This conservation of water on such a great scale offers striking contrast to the truly prodigal habits of certain plants that actually drip water, so charged are they with this precious liquid, and so little stress do their conditions of life put them under in this respect. Where water is plentiful and turgor maintained almost to the bursting point, evaporation in a moist or chilly atmosphere does not suck out water vapor fast enough. Sometimes, around the edges of the leaves of the common garden nasturtium, drops of water may be found, literally forced out as drops, rather than transpired as water vapor. This happens to a considerable number of plants, during the night when transpiration is laggard, and such drops are usually mistaken for dew. The latter is actually the condensation of moisture in the air upon the leaves of plants which cool down more rapidly than the air, and seldom due to the forcing out of drops of water from leaves, although in rare cases it may be. In tropical forests, where the humidity is very heavy and water supply from the roots copious, certain leaves leak water so fast and are so constructed that this excess is prevented from accumulating on the leaf. The pipal tree of India has long drip tips to its leaves that conduct the excess water from the blade to the end of the slender tip where it drips off. The advantage of these dripping points is obvious, for in regions so humid that water is forced out of the leaf, the coating of the leaf with this extra moisture would by that much retard transpiration. Dripping points, which in less exaggerated forms than in the pipal tree are common in many parts of the world, are thus of decided advantage.
Whether it be desirous to retain water or to lose it by gradual evaporation, or expel an excess of it, each species of plant has developed the apparatus to best preserve its individual life. While only the barest outline of these adjustments to the water requirements of plants has been given here, the details form an almost dramatic picture of struggle of the different kinds of plants for survival. The extremes are the desert plants on the one hand and those of the rain forests in the tropics on the other. The chapter on Plant Distribution will show how important these water requirements of plants have been in determining what grows on the earth to-day.
With carbon dioxide going in, oxygen, water vapor and, as we have seen, even liquid water coming out of the stoma of leaves, it might be surmised that these busy little pores and their guard cells had done work enough for the plant. And yet there is still one more act to play and the stoma have much to do with it. For this process of photosynthesis and the closely related one of supplying food and water to the leaf cannot go on without respiration, which is quite another thing. In plants respiration or breathing has no more to do with digestion than it does in man. Digestion in man is not unlike photosynthesis in plants, except that plants make food in the process while men destroy it. But plants must breathe just as we do, and, as we need oxygen to renew our vital processes, so do they. While respiration is a necessary part of plant activity it is not such an important part as photosynthesis, for which it is often mistaken. The thing to fix in our minds is that photosynthesis makes food, uses the sun’s energy and releases oxygen in the process, while respiration uses oxygen and might almost be likened to the oil of a machine—necessary but producing nothing.
5. Restless and Irritable Plants
In walking through the quiet cathedrallike stillness of a deep forest or over the fields and moors, perhaps our chief thought is how restful the scene is, and what a contrast the quiet, patient plants make to the darting insects or flitting birds that our walk disturbs. We found at the beginning of this book that ability to get about is one of the main differences between animals and plants. Like so many first thoughts, this is, however, only a half truth, for while most plants, seemingly by a kind of fatality, are anchored forever to the place of their birth, many of them do move certain parts of themselves and that quite regularly. While some of these movements have already been hinted at as a possible response to transpiration or too intense light, there are others where the advantage to the plant, if any, has yet to be demonstrated. These other movements, perhaps because their cause has never been discovered, seem the more mysterious as they certainly are more weird and interesting than almost any other of the curious things that plants do.
Perhaps the most difficult thing in the world is to keep an active growing child perfectly still for more than a few moments at a time. There seems to be some impelling force that makes young growing things in a constant state of restlessness, and it is perhaps not so extraordinary, after all, that practically all young plants are restless in the sense that they are never quite still. And, like many grown-up people who do not know what repose in their waking moments really means, there are a goodly number of plants that are restless until the day they die.
Charles Darwin, perhaps the greatest man that the last century produced, wrote a book in two volumes on these restless plants, and proved by a series of experiments illustrated by charts which the plants themselves drew for him, that there were perhaps no plants that do not move at least some part of themselves during the early stages of their career. While he never could explain the cause of these movements he left in that book an imperishable record of the amount and direction of these mysterious movements, which are almost to be likened to the growing pains of young children.
The tips or growing shoots of many plants will point in one direction in the early morning, a different way at noon and still a different one by nightfall. Hundreds of totally unrelated plants seem to have this habit of moving their tips through a definite cycle during each day and this restlessness does not appear to be of the slightest use to them. It cannot be response to the moving of the sun through the sky, for often the movement may be away from the direct sunshine, and sometimes the motion goes on in the dark, as experiments have proved.
It is hard to see the movement of the whole upper part of a plant, although it is well known that they do move in many cases. But in the tendrils the movement is often easy to observe and even to induce. Some of these slender aids to climbing plants, if they happen to be swinging freely in the air, do actually make slow circular movements, that even if they were designed for the purpose could not more perfectly accomplish their obvious intent, which is to catch the nearest favorable support. These circular movements are to the left in the hop, honeysuckle and many other plants, to the right in the climbing beans, morning-glory and some others. When the tendril reaches a support it almost immediately turns about it, in the same direction as its free movements through the air have been. It is thus this apparently aimless swinging of tendrils through space that determines whether the vine is going to twine to the right or left. The speed with which a tendril will take its first turns about a support is so comparatively rapid that, once the support is caught there is scarcely a chance of the vine being torn away by the wind or other agency as would surely happen if tendril movements were the leisurely things that some folks think they are. In the case of one Passion-flower vine, which are gorgeous climbers mostly from the tropics, the tendril made a complete turn in two minutes after it first touched a possible support. And there is a quite noticeable movement in thirty seconds if the tip of the tendril be ever so lightly touched. Teasing tendrils to see how much or how fast they will coil has resulted in some extraordinary cases of the “comeback” of some of them. Darwin irritated a tendril for a few moments and induced a partial coiling which straightened out when the object causing it was withdrawn. To see how long the plant would stand this sort of thing and still not be literally tired of coiling he succeeded in making the plant partially coil, and by withdrawing the incentive uncoil again, over twenty times in fifty-four hours. An impulse to coil of such persistence as this naturally results in vines forming the impenetrable thickets they do in many forests. It emphasizes how restless are the growing points of these climbers, and serves as a striking illustration of those gradual movements of many other plants that seem to have some relation to growth, but in a way not yet understood. For while it is an obvious advantage for the vine to swing its tendrils through the air this advantage has not yet been proved the cause of the swinging. In fact if all possible supports are removed the tendril will often coil anyway, a perfectly futile proceeding, that looks almost like disgust.
This general restlessness, which by the imaginative has been thought of as a mild protest by plants at their otherwise fixed position, is not so spectacular as that of certain other plants, notably the poplars. A flattened instead of a round leafstalk makes the leaves of these trees flutter in the lightest air and in a gale the tree is a mass of animated foliage. No use has ever been found for this curious habit and it is not certain that it is of the least advantage to the tree. If anything, the constant movement may have the decided disadvantage of increasing transpiration.
In our common wood sorrel the leaflets on cloudy days or during the night regularly “go to sleep.” That is, they are folded at such times, rather than spread out in the ordinary way. These sleep movements may have something to do with transpiration, but whether or not this is true they are very regular and in certain plants the habit is remarkably and rather mysteriously uniform. Why, for instance, do the leaflets of these wood sorrels, the beans, lupine, locust tree and licorice plant, always fold downward while the clovers, vetch, peas, and bird’s-foot trefoil are always folded upward? Such movements and their direction are among the unsolved problems of botany, and merely to know of them or observe them leads us nowhere as to their true inwardness.
But quite apart from these merely restless plants, and there are thousands of different kinds which are known to move slightly, at least during their young stages, are a few more decidedly active ones that are seemingly irritable. At least they show peculiar movements if touched, and at night. One of the best known is the sensitive plant from tropical America. Its twice compound leaf is composed of many tiny leaflets which upon the slightest touch close up and apparently wither on their stalk at once. In five seconds after the leaf is touched it will appear like a wilted wreck. If the jar is sharp enough the whole plant will droop, and the response to a sudden jar is almost electrically quick in its action. And yet all this sudden wilting, actually caused by a quick loss of turgor, is slowly repaired and the plant carries on quite normally again until another shock renews its irritable response. This plant does the same thing gradually during the night, except that the leaflets recover their normal position only with the rise of the sun.
From India comes the most remarkable of all plants so far as movements are concerned. For in the telegraph plant the movements are so regular and long continued that irritability might almost be said to be continuous. The plant is a low shrub or herb with compound leaves, and the terminal leaflet, which is much larger than its neighbors on either side of their common stalk, performs a motion that describes with its tip an irregular oval or ellipse. But the movement is not steady; it goes by a series of slight but perfectly distinct jerks. It takes about two minutes for the leaf to complete its cycle, and it is this jerky movement that has given the plant its name. During the night its leaflets stop this apparently quite useless performance, the cause of which is quite unknown. It is often grown in greenhouse collections where its strange movements may be seen on any sunny day.
Many other cases of the restlessness or irritability of plants could be given, and nothing has been said here of the curious movements of some insectivorous plants as they have already been mentioned. The very considerable movements of certain flower and fruit organs will also be considered elsewhere.
It cannot have escaped the thoughtful reader that all of this chapter on plant behavior has dealt with those functions of plants in which roots, stems, or leaves play the chief part. These purely vegetative actions of plants, what might almost be called their bread and butter activities, would never lead to perpetuating their kind. For while all of these functions are necessary, except certain apparently wayward movements which still remain unexplained, they are in a sense only the preparation for an infinitely more important act, the reproduction of their kind. What the poetic have called the love of the flowers, or in more prosaic but perhaps more truthful words the fertilization, pregnancy, and birth of the new race, will be considered in a separate chapter. No other act of the plant world is so interesting as the mechanism of reproduction, the almost endless devices for securing it, and the ingenuity of nature in seeing to it that there are no flukes.